
Fusaricidins, Polymyxins and Volatiles Produced by Paenibacillus polymyxa Strains DSM 32871 and M1

 ,
,
Abstract
:1. Introduction
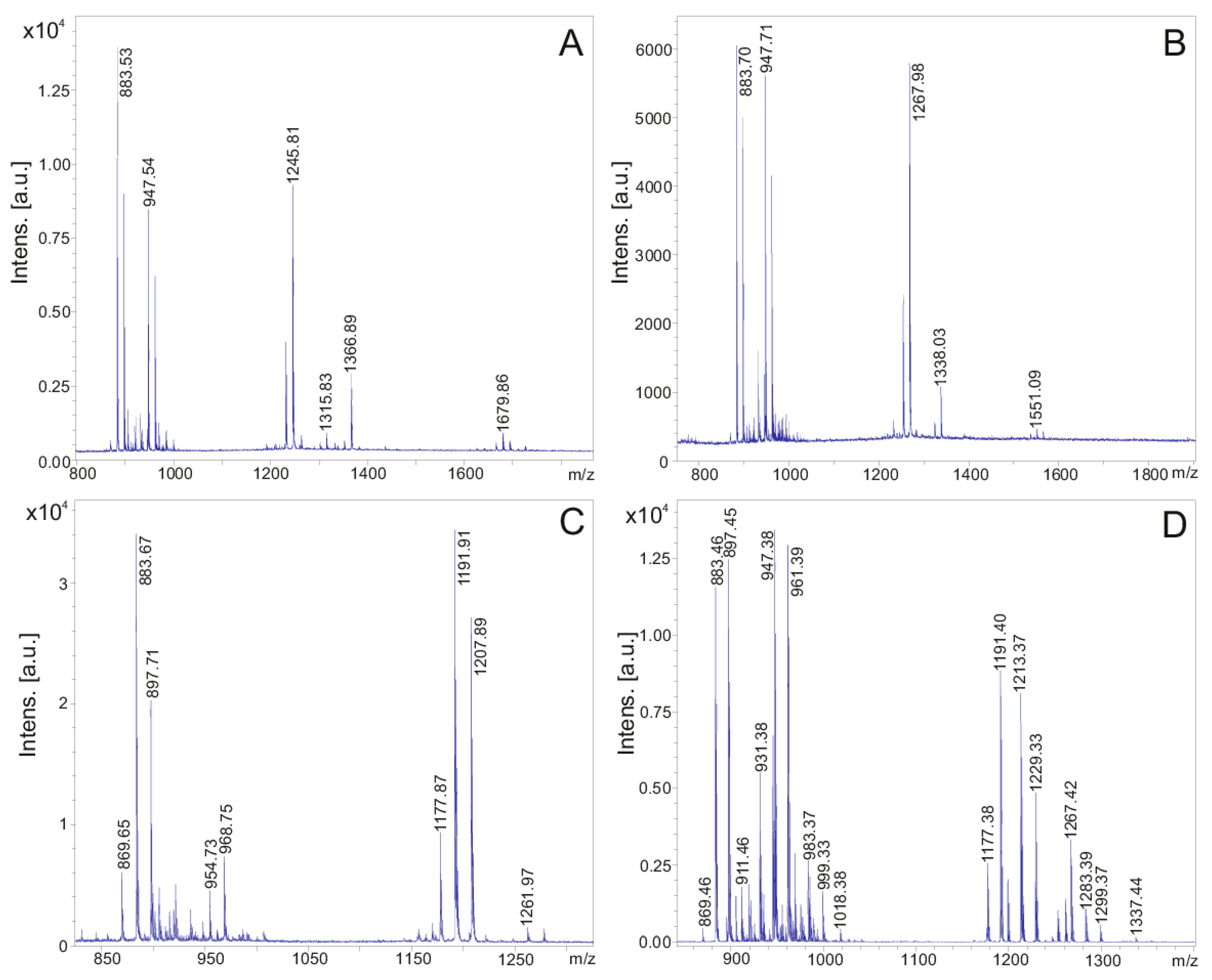
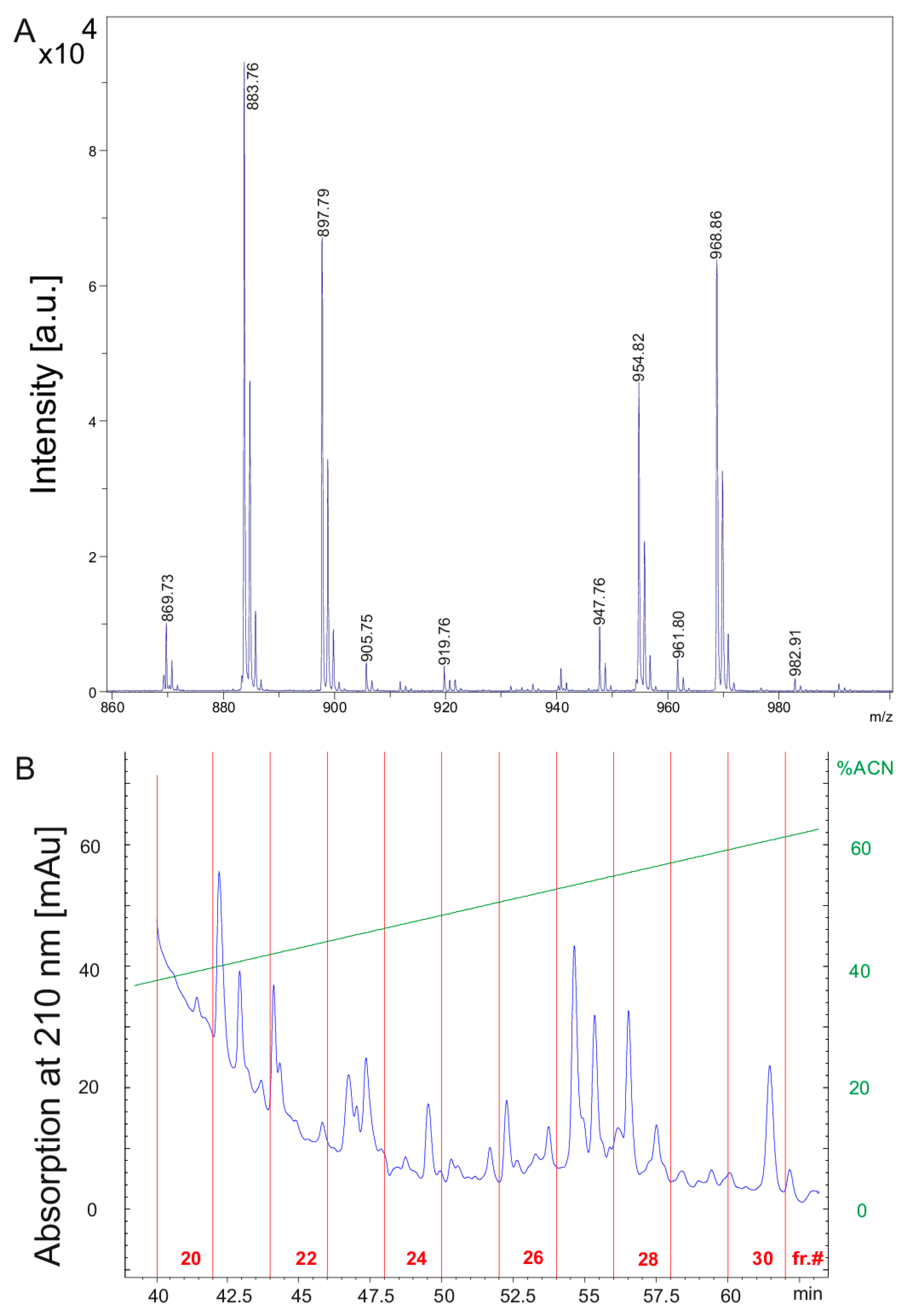
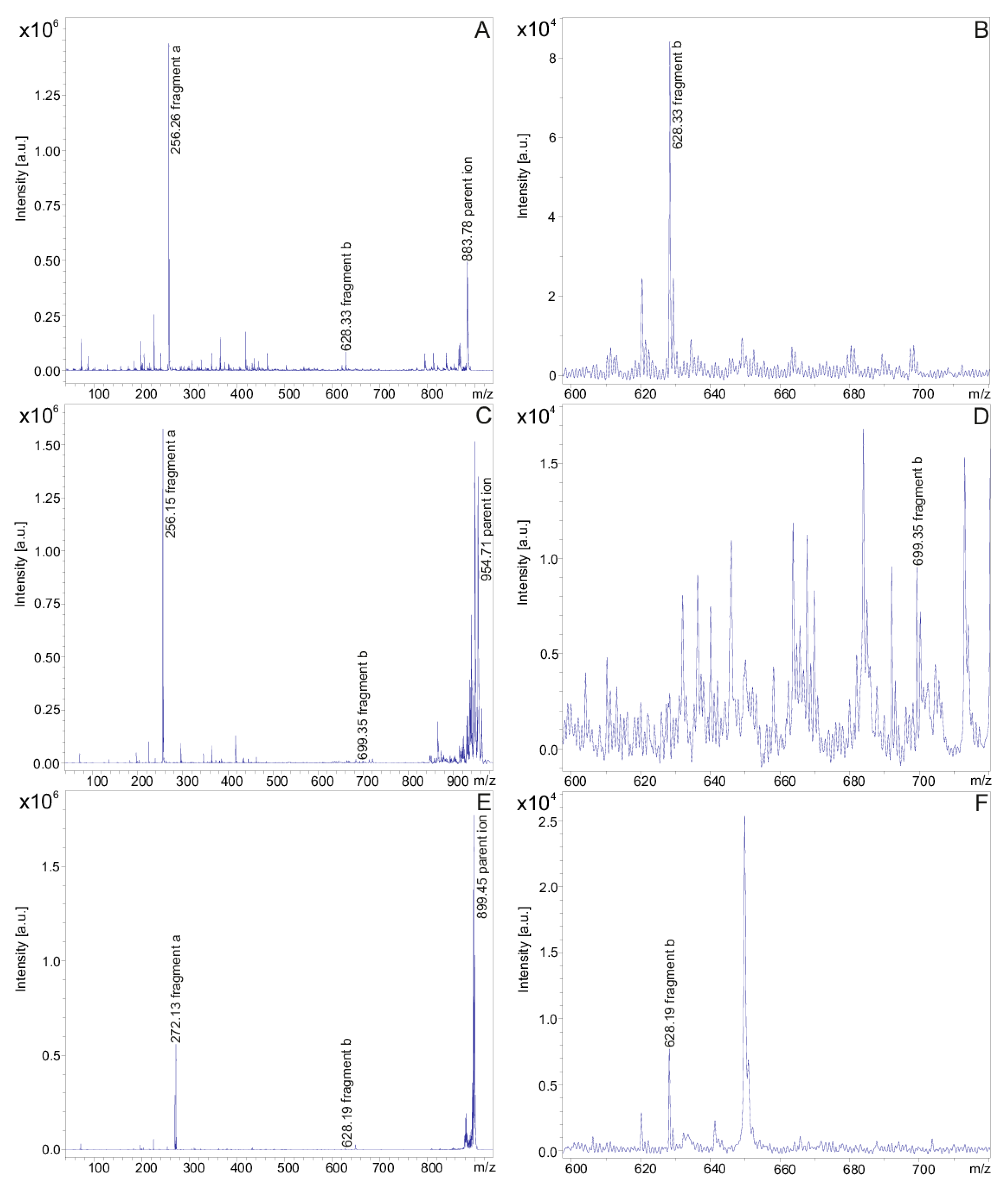
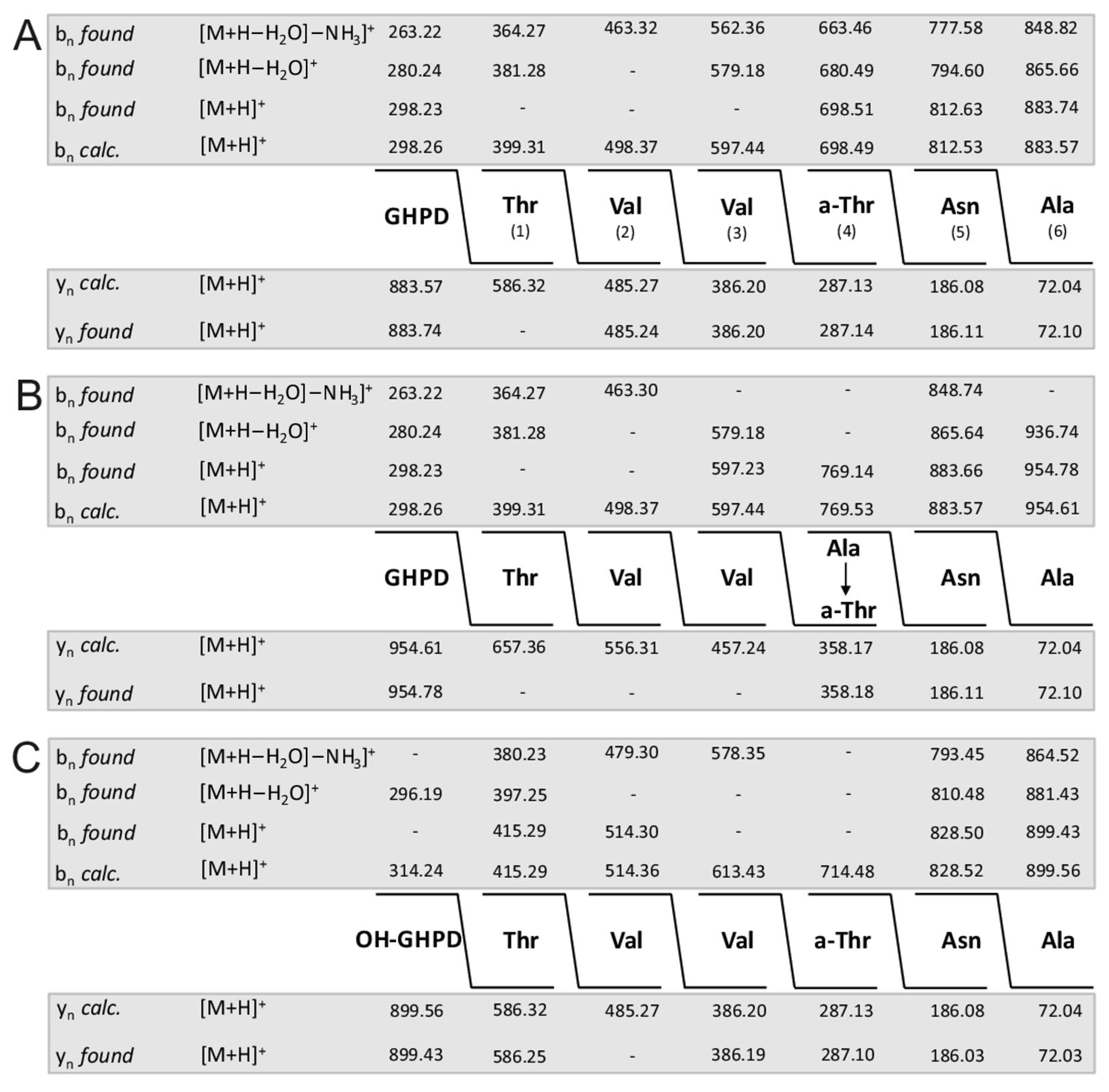
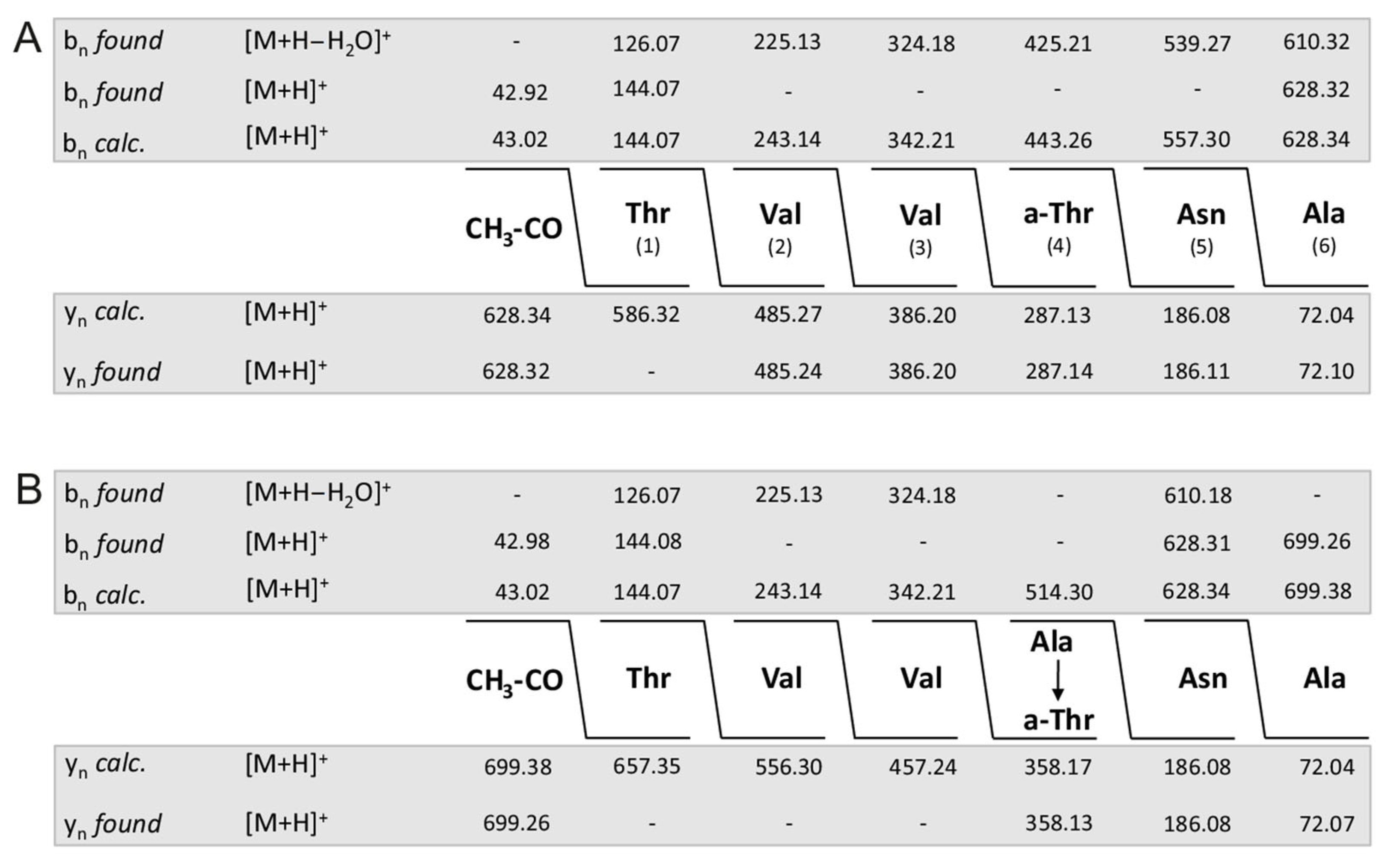
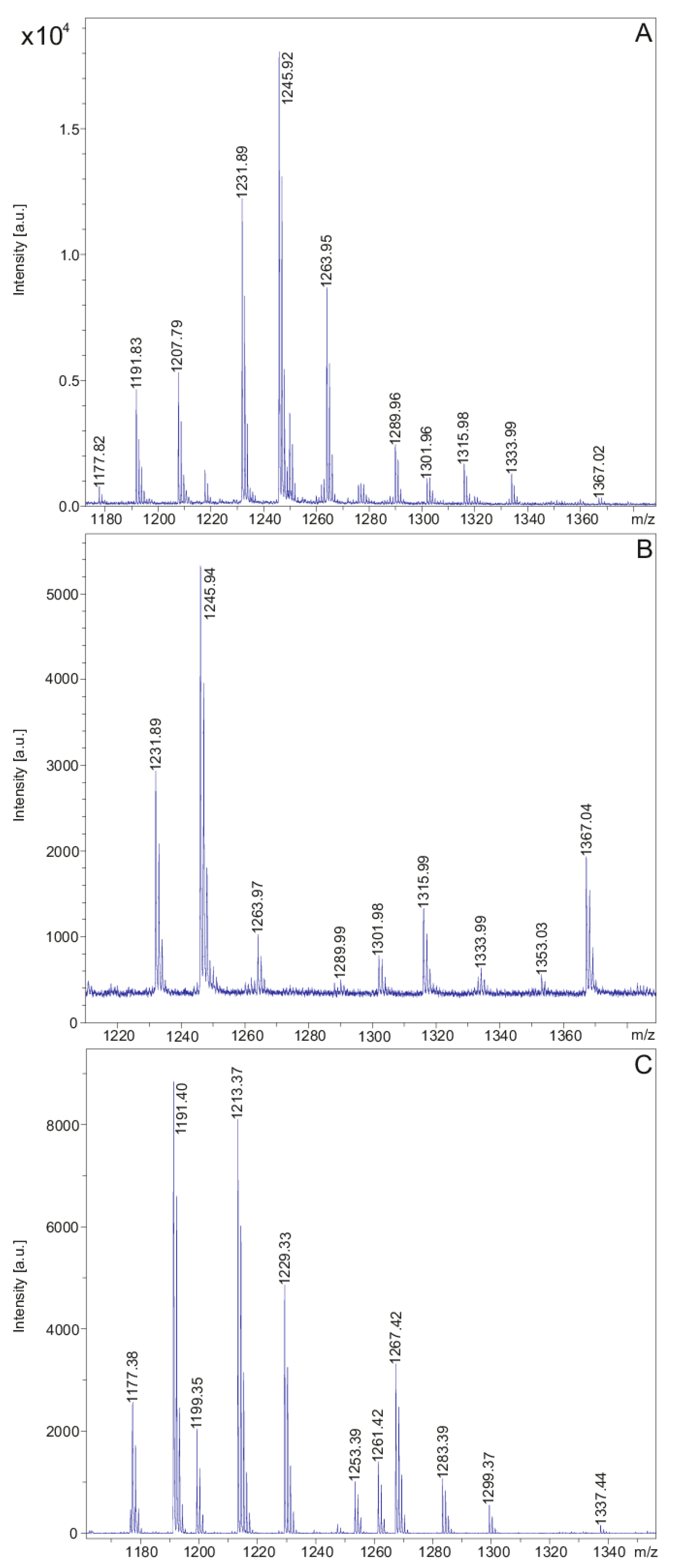
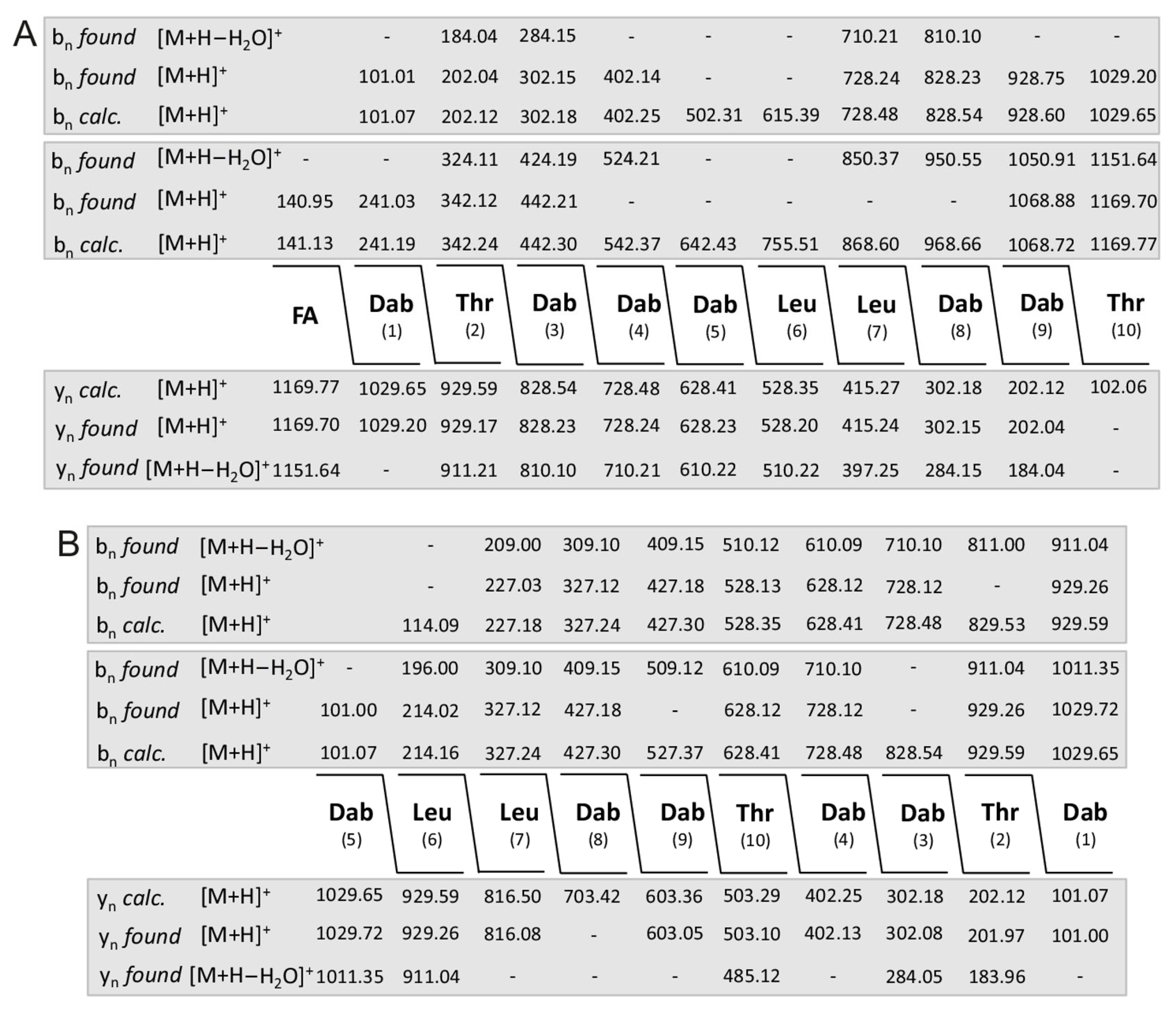
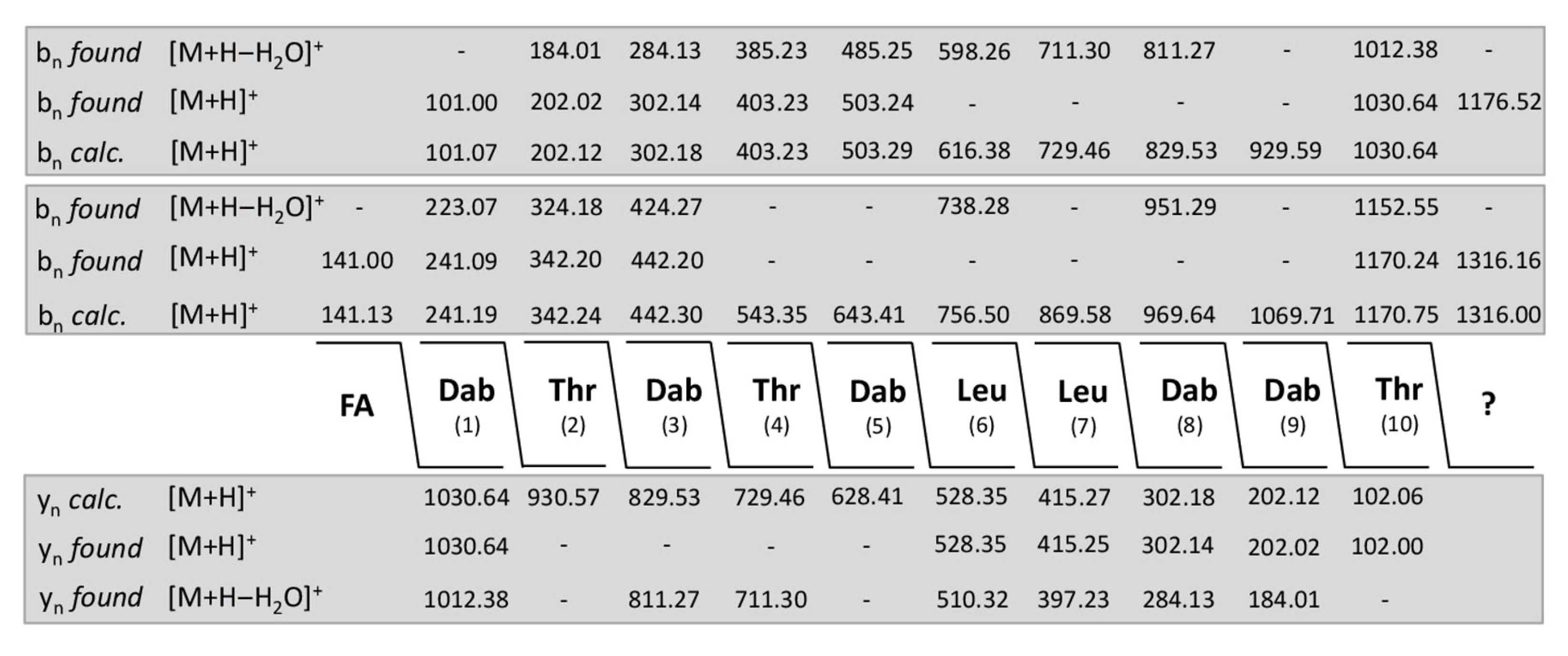
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cultivation of Organisms
4.3. Sample Preparation
4.4. HPLC Fractionation of the Bioactive Compounds
4.5. Profiling of Bioactive Peptides by MALDI-TOF MS
4.6. Identification of Microbial Volatiles Using GC–MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus spp.: A gold mine of antibiotic candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Aleti, G.; Sessitsch, A.; Brader, G. Genome mining: Prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Comput. Struct. Biotechnol. 2015, 13, 192–203. [Google Scholar] [CrossRef]
- Xie, J.; Shi, H.; Du, Z.; Wang, T.; Liu, X.; Chen, S. Comparative genomic and functional analysis reveal conservation of plant growth promoting traits in Paenibacillus polymyxa and its closely related species. Sci. Rep. 2016, 6, 21329. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajimura, Y.; Kameda, M. Fusaricidin A, a new depsipeptide antibiotic produced by Bacillus polymyxa KT-8. Taxonomy, fermentation, isolation, structure elucidation and biological activity. J. Antibiot. 1996, 49, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Kajimura, Y.; Kameda, M. Fusaricidin B, C and D, new depsipeptide antibiotics produced by Bacillus polymyxa KT-8. Isolation, structure elucidation and biological activity. J. Antibiot. 1997, 50, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Kurusu, K.; Ohba, K.; Arai, T.; Fukushima, K. New peptide antibiotics LI-F03, F04, F05, F07 and F08 produced by Bacillus polymyxa. Isolation and characterization. J. Antibiot. 1997, 40, 1506–1514. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, J.; Fukai, J.; Konishi, M.; Ono, J.; Kurusu, K.; Nomura, T. Li-F antibiotics, family of antifungal cyclic depsipeptides produced by Bacillus polymyxa L-1129. Heterocycles 2000, 53, 1533–1549. [Google Scholar] [CrossRef]
- Kuroda, J.; Fukai, J.; Nomura, T. Collision-induced dissociation of ring-opened cyclic depsipeptides with a guanidino group by electrospray ionization trap mass spectrometry. J. Mass Spectrom. 2001, 36, 30–37. [Google Scholar] [CrossRef]
- Vater, J.; Niu, B.; Dietel, K.; Borriss, R. Characterization of novel fusaricidins produced by Paenibacillus polymyxa-M1 using MALDI-TOF mass spectrometry. J. Am. Soc. Mass Spectrom. 2015, 26, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Vater, J.; Herfort, S.; Doellinger, J.; Weydmann, M.; Dietel, K.; Faetke, S.; Lasch, P. Fusaricidins from Paenibacillus polymyxa M-1, a family of lipohexapeptides of unusual complexity—A mass spectrometric study. J. Mass Spectrom. 2017, 52, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Avula, B.; Guan, S.; Ravu, R.R.; Wang, M.; Zhao, J.; Khan, I.A.; Hinchee, M.; Li, X.-C. Identification of fusaricidins from the antifungal microbial strain Paenibacillus sp. MS2379 using ultra-high performance liquid chromatography coupled to quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2019, 1586, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Vater, J.; Herfort, S.; Doellinger, J.; Weydmann, M.; Borriss, R.; Lasch, J. Genome mining of the lipopeptide biosynthesis of Paenibacillus polymyxa E681 in combination with mass spectrometry: Discovery of the lipoheptapeptide paeniloheptin. ChemBioChem 2018, 19, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Roberts, K.D.; Li, J. Rediscovering the octapeptins. Nat. Prod. Rep. 2017, 34, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Sogn, J.A. Structure of the peptide antibiotic polypeptin. J. Med. Chem. 1976, 19, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.-D.; Liu, T.-Z.; Zhou, S.-L.; Ding, R.; Zhao, W.-P.; Li, O.; Wu, X.-C. Identification and functional analysis of gene cluster involvement in biosynthesis of the cyclic lipopeptide antibiotic pelgipeptin produced by Paenibacillus elgii. BMC Microbiol. 2012, 12, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure-activity relationships of polymxin antibiotics. J. Med. Chem. 2010, 53, 1898–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Hinoo, H.; Shoji, J. The structure of tridecaptin A (studies on antibiotics from the genus Bacillus. XXIV). J. Antibiot. 1978, 31, 652–661. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Sakazaki, R.; Hinoo, H.; Shoji, J. The stuctures of tridecaptins B and C (studies on antibiotics from the genus Bacillus. XXV). J. Antibiot. 1979, 32, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, S.A.; Lohans, C.T.; van Belkum, M.J.; Bels, M.A.; Vederas, J.C. Studies on tridecapeptin B(1), a lipopeptide with activity against multidrug resistant Gram-negative bacteria. Org. Biomol. Chem. 2015, 13, 6073–6081. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Huang, E.; Yuan, C.; Zhang, L.; Yousef, A.E. Isolation of a Paenibacillus sp. strain and structural elucidation of its broad-spectrum lipopeptide antibiotic. Appl. Environ. Microbiol. 2012, 78, 3156–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, E.; Guo, Y.; Yousef, A.E. Biosynthesis of the new broad-spectrum lipopeptide antibiotic paenibacterin in Paenibacillus thiaminolyticus OSY-SE. Res. Microbiol. 2014, 165, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Lal, S.; Tabacchioni, S. Ecology and biotechnological potential of Paenibacillus polymyxa: A minireview. Ind. J. Microbiol. 2009, 49, 2–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, A.W.; Heinrichs, D.E.; Yuan, Z.C. Comparative and genetic analysis of four sequenced Paenibacillus polymyxa genomes reveals a diverse metabolism and conservation of genes relevant to plant-growth promotion and competitiveness. BMC Genom. 2014, 15, 851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.A.; Lou, Y.; Hafeez, R.; Li, X.; Hossein, A.; Xie, T.; Lin, L.; Li, B.; Yin, Y.; Yan, J.; et al. Functional analysis of genome mining reveal high potential of biocontrol and plant growth promotion in nodule-inhabiting bacteria within Paenibacillus polymyxa complex. Front. Microbiol. 2021, 11, 618601. [Google Scholar] [CrossRef] [PubMed]
- Mousa, W.K.; Shearer, C.R.; Limay-Rios, V.; Zhou, T.; Raizada, M.N. Bacterial endophytes from wild maize suppress Fusarium graminearum in modern maize and inhibit mycotoxin accumulation. Front. Plant Sci. 2015, 6, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. Isolation and characterization of plant growth-promoting endophytic bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. BioMed Res. Int. 2020, 2020, 8650957. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Rueckert, C.; Blom, J.; Wang, Q.; Borriss, R. The genome of the plant growth promoting rhizobacterium Paenibacillus polymyxa M-1 contains nine sites dedicated for nonribosomal synthesis of lipopeptides and polyketides. J. Bacteriol. 2011, 193, 5862–5863. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S. Fusaricidin produced by Paenibacillus polymyxa WLY78 induces systemic resistance against Fusarium wilt of cucumber. Int. J. Mol. Sci. 2019, 20, 5240. [Google Scholar] [CrossRef] [Green Version]
- Beatty, P.H.; Jensen, S.E. Paenibacillus polymyxa produces fusaricidin-type antifungal antibiotics active against Leptoshaeria maculans, the causative agent of blackleg disease of canola. Can. J. Microbiol. 2002, 48, 159–169. [Google Scholar] [CrossRef]
- Abdallah, Y.; Yang, M.; Zhang, M.; Masum, M.M.I.; Ogunyemi, S.O.; Hossein, A.; An, Q.; Yan, C.; Li, B. Plant growth promotion and suppression of bacterial leaf blight in rice by Paenibacillus polymyxa Sx3. Lett. Appl. Microbiol. 2019, 68, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Roberts, K.D.; Nation, R.L.; Wang, J.; Thompson, P.E.; Li, J. Teaching old polymyxins new tricks: New-generation lipopeptides targeting Gram-negative superbugs. ACS Chem. Biol. 2014, 9, 1172–1177. [Google Scholar] [CrossRef]
- Suzuki, T.; Inouye, H.; Fujikawa, K.; Suketa, Y. Studies on the chemical structure of colistin. J. Biochem. 1963, 54, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hayashi, K.; Fujikama, K.; Tsukamoto, K. The chemical structure of polymyxin E: The identities of polymyxin E1 with colistin A and of polymyxin E2 with colistin B. J. Biochem. 1965, 57, 226–227. [Google Scholar] [CrossRef]
- Govaerts, C.; Rozenski, J.; Orwa, J.; Roets, E.; Van Schepdael, A.; Hoogmartens, J. Mass spectrometric fragmentation of cyclic peptides belonging to the polymyxin and colistin antibiotics studied by ion trap and quadrupole/orthogonal-acceleration time-of-flight technology. Rapid Commun. Mass Spectrom. 2002, 16, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Murai, E.; Fujisawa, M.; Tatsuki, T.; Nobue, F. Polymyxin P, new antibiotic of polymyxin group. J. Antibiot. 1969, 22, 449–450. [Google Scholar] [CrossRef]
- Brown, P.; Dawson, M.J. Development of new polymyxin derivatives for multi-drug resistant Gram-negative infections. J. Antibiot. 2017, 70, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govaerts, C.; Orwa, J.; Van Schepdael, A.; Roets, E.; Hoogmartens, J. Characterization of the polymyxin series by liquid chromatography electrospray ionization ion trap tandem mass spectrometry. J. Pept. Sci. 2002, 7, 45–55. [Google Scholar] [CrossRef]
- Govaerts, C.; Orwa, J.; Van Schepdael, A.; Roets, E.; Hoogmartens, J. Liquid chromatography-ion trap tandem 39mamass spectrometry for the characterization of polypeptide antibiotics of the colistin series in commercial samples. J. Chromatogr. A 2002, 976, 65–78. [Google Scholar] [CrossRef]
- Orwa, J.; Govaerts, C.; Gevers, K.; Roets, E.; Van Schepdael, A.; Hoogmartens, J. Study of the stability of polymyxins B1, E1 and E2 in aqueous solution using liquid chromatography and mass spectrometry. J. Pharm. Biomed. Anal. 2002, 29, 203–212. [Google Scholar] [CrossRef]
- Shaheen, M.; Li, J.; Ross, A.C.; Vederas, J.C.; Jensen, S.E. Paenibacillus polymyxa PK31 produces variants of polymyxin B-type antibiotics. Chem. Biol. 2011, 18, 1640–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambadou, F.; Caradec, T.; Gagez, A.-L.; Bonnet, A.; Sopena, V.; Bridiau, N.; Thiéry, V.; Didelot, S.; Barthélémy, C.; Chevrot, R. Characterization of the colistin (polymyxin E1 andE2) biosynthetic gene cluster. Arch. Microbiol. 2015, 197, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-K.; Park, S.-Y.; Kim, R.; Kim, S.-B.; Lee, C.-H.; Kim, J.F.; Park, S.-H. Identification of a polymyxin synthetase gene cluster of Paenibacillus polymyxa and heterologous expression of the gene in Bacillus subtilis. J. Bacteriol. 2009, 191, 3350–3358. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Vater, J.; Rueckert, C.; Blom, J.; Lehmann, M.; Ru, J.-J.; Chen, X.-H.; Wang, Q.; Borriss, R. Polymyxin P is the active principle in suppressing phytopathogenic Erwinia spp. by the biocontrol rhizobacterium Paenibacillus polymyxa M-1. BMC Microbiol. 2013, 13, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaara, M. Polymyxins and their potential next generation as therapeutic antibiotics. Front. Microbiol. 2019, 10, 1689. [Google Scholar] [CrossRef] [PubMed]
- Nang, S.C.; Azad, M.A.K.; Velkov, T.; Zhou, Q.; Li, J. Rescuing the last-line polymyxins: Achievements and challenges. Pharmacol. Rev. 2021, 73, 679–728. [Google Scholar] [CrossRef]
- Audrain, B.; Farag, M.A.; Ryu, C.M.; Ghigo, J.M. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanschiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Kai, M. Diversity and distribution of volatile secondary metabolites throughout Bacillus subtilis isolates. Front. Microbiol. 2020, 11, 559. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Pare, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Asari, S.; Matzen, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple effects of Bacillus amyloliquefaciens volatile compounds: Plant growth promotion and growth inhibition of phytopathogens. FEMS Microbiol. Ecol. 2016, 92, 070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. Microbiol. Open 2018, 8, e813. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Klöpper, J.W.; Pare, P.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, M.A.; Ryu, C.-M.; Sumner, L.W.; Pare, P.W. GC-MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar] [CrossRef]
- Suckau, D.; Resemann, A.; Schuerenberg, M.; Hufnagel, P.; Franzen, J.; Holle, A. A novel MALDI LIFT-TOF/TOF mass spectrometer for proteomics. Anal. Bioanal. Chem. 2003, 376, 952–965. [Google Scholar] [CrossRef]
- Mülner, P.; Schwarz, E.; Dietel, K.; Junge, H.; Herfort, S.; Weydmann, M.; Lasch, P.; Cernava, T.; Berg, G.; Vater, J. Profiling for bioactive peptides and volatiles of plant growth promoting strains of the Bacillus subtilis complex of industrial relevance. Front. Microbiol. 2020, 11, 1432. [Google Scholar] [CrossRef]
- Ferreira, S.B.; Kaiser, C.R. Pyrazine derivatives: A patent review (2008-present). Expert Opin. Ther. Pat. 2012, 22, 1033–1151. [Google Scholar] [CrossRef]
- Haidar, R.; Roudet, J.; Bonnard, O.; Dufour, M.C.; Corio-Costet, M.F.; Fert, M.; Gautier, P.; Deschamps, A.; Fermaud, M. Screening and modes of action of antagonistic bacteria to control the fungal pathogen Phaeomoniella chlamydospora involved in grapevine trunk diseases. Microbiol. Res. 2016, 192, 172–184. [Google Scholar] [CrossRef]
- Kusstatscher, P.; Cernava, T.; Liebminger, S.; Berg, G. Replacing conventional decontamination of hatching eggs with a natural defense strategy based on antimicrobial volatile pyrazines. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mülner, P.; Wagner, P.; Bergna, A.; Sarajlic, D.; Gstöttenmayr, B.; Dietel, K.; Grosch, R.; Cernava, T.; Berg, G. Microbiota associated with sclerotia of soilborne fungal pathogens—A novel source of biocontrol agents producing bioactive volatiles. Phytobiom. J. 2019, 3, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Fernando, W.G.D.; Ramarathnam, R.; Krishnamoorthy, A.S.; Savchuk, S.C. Identification and use of potential bacterial organic antifungal volatiles in biocontrol. Soil Biol. Biochem. 2005, 37, 955–964. [Google Scholar] [CrossRef]
- Popova, A.A.; Koksharova, O.A.; Lipasova, V.A.; Zaitseva, J.V.; Katkova-Zhukotskaya, O.A.; Eremina, S.I.; Mironov, A.S.; Chernin, L.S.; Khmel, I.A. Inhibitory and toxic effects of volatiles emitted by strains of Pseudomonas and Serratia on growth and survival of selected microorganims, Caenorhabditis elegans and Drosophila melanogaster. Biomed. Res. Int. 2014, 2014, 125704. [Google Scholar] [CrossRef] [Green Version]
- Toffano, L.; Fialho, M.B.; Pascholati, S.F. Potential of fumigation of orange fruits with volatile organic compounds produced by Saccharomyces cerevisiae to control citrus black spot disease at postharvest. Biol. Control. 2017, 108, 77–82. [Google Scholar] [CrossRef]
- Fincheira, P.; Venthur, H.; Mutis, A.; Parada, M.; Quirot, A. Growth promotion of Lactuca sativa in response to volatile organic compounds emitted from diverse bacterial species. Microbiol. Res. 2016, 193, 39–47. [Google Scholar] [CrossRef]
- Han, S.H.; Lee, S.J.; Moon, J.H.; Park, K.H.; Yang, K.Y.; Cho, B.H.; Kim, K.Y.; Kim, Y.W.; Lee, M.C.; Anderson, A.J.; et al. GacS-dependent production of 2R, 3R butanediol by Pseudomonas chlororaphis 06 is a major determinant for eliciting resistance against Erwinia carotovora but not against Pseudomonas syringae pv. Tabaci. Mol. Plant Microbe Interact. 2006, 19, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Ledger, T.; Rojas, S.; Timmermann, T.; Pinedo, I.; Poupin, M.J.; Garrido, T.; Richter, P.; Tamayo, J.; Donoso, R. Volatile-mediated effects predominate in Paraburkholderia phytofirmans growth promotion and salt stress tolerance of Arabidopsis thaliana. Front. Microbiol. 2016, 7, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudrappa, T.; Biedrzycki, M.L.; Kunjeti, S.G.; Donofrio, N.M.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. The rhizobacterial elicitor acetoin induces systemic resistance in Arabidopsis thaliana. Commun. Integr. Biol. 2010, 3, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okonkwo, C.C.; Ujor, V.; Cornish, K.; Ezeji, T.C. Inactivation of the levansucrase gene in Paenibacillus polymyxa DSM 365 diminishes exopolysaccharide biosynthesis during 2,3-butanediol fermentation. Appl. Environ. Microbiol. 2020, 86, e00196-20. [Google Scholar] [CrossRef]
- Lo Cantore, P.; Giorgio, A.; Iacobellis, N.S. Bioactivity of volatile organic compounds produced by Pseudomonas tolaasii. Front. Microbiol. 2015, 6, 1082. [Google Scholar] [CrossRef] [Green Version]
- Stotzky, G.; Schenk, S. Volatile organic compounds and microorganisms. CRC Crit. Rev. Microbiol. 1976, 4, 333–382. [Google Scholar] [CrossRef]
- Hertel, M.; Preissner, R.; Gilissen, B.; Schmidt-Westhausen, A.M.; Paris, S.; Preissner, S. Detection of signature volatiles for cariogenic microorganisms. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Splivallo, R.; Novero, M.; Bertea, C.M.; Bossi, S.; Bonfante, P. Truffle volatiles inhibit growth and induce an oxidative burst in Arabidopsis thaliana. New Phytol. 2007, 175, 417–424. [Google Scholar] [CrossRef]
- Weise, T.; Kai, M.; Gummesson, A.; Troeger, A.; von Reuß, S.; Piepenborn, S.; Kosterka, F.; Sklorz, M.; Zimmermann, R.; Francke, W.; et al. Volatile organic compounds produced by the phytopathogenic bacterium Xanthomonas campestris pv. vesicatoria 85-10. Beilstein J. Org. Chem. 2012, 8, 579–596. [Google Scholar] [CrossRef] [Green Version]
- Lemfack, M.C.; Gohlke, B.-O.; Toguem, S.M.; Preissner, S.; Piechulla, B.; Preissner, R. mVOC 2.0: A database of microbial volatiles. Nucleic Acids Res. 2018, 46, D1261–D1265. [Google Scholar] [CrossRef] [Green Version]
- Stansly, P.G.; Schlosser, M.E. Studies on polymyxin: Isolation and identification of Bacillus polymyxa and differentiation of polymyxin from certain known antibiotics. J. Bacteriol. 1947, 54, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Rybakova, D.; Rack-Wetzlinger, U.; Cernava, T.; Schaefer, A.; Schmuck, M.; Berg, G. Aerial warfare: A volatile dialogue between the plant pathogen Verticillium longisporum and its antagonist Paenibacillus polymyxa. Front. Plant Sci. 2017, 8, 1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C.-M. Induced resistance by a long-chain bacterial volatile: Elicitation of plant systemic defense by a C13 volatile produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef] [PubMed]
- Yegorenkova, I.V.; Tregubova, K.V.; Krasov, A.I.; Evseeva, N.V.; Matora, L.Y. Effect of exopolysaccarides of Paenibacillus polymyxa rhizobacteria on physiological and morphological variables of wheat seedlings. J. Microbiol. 2021, 59, 729–735. [Google Scholar] [CrossRef]
- He, X.; Li, Q.; Wang, N.; Chen, S. Effects of an EPS biosynthesis gene cluster of Paenibacillus polymyxa WLY78 on biofilm formation and nitrogen fixation under aerobic conditions. Microorganisms 2021, 9, 289. [Google Scholar] [CrossRef] [PubMed]
- Landy, M.; Warren, G.H.; Rosenman, S.B.; Colio, L.G. Bacillomycin; an antibiotic from Bacillus subtilis active against pathogenic fungi. Proc. Soc. Exp. Biol. Med. 1948, 67, 539–541. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
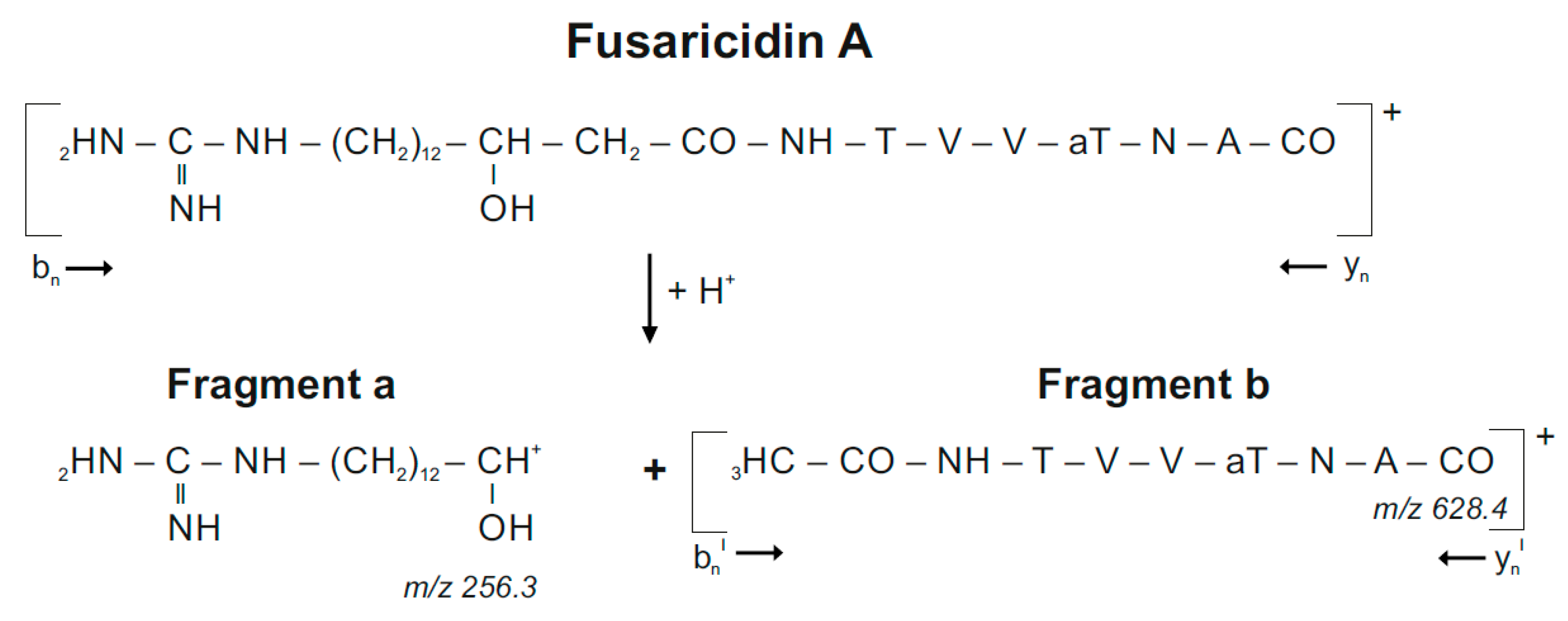{kind=link}
| Fusaricidins m/z | m/z + 71 | Fragment a m/z | Fragment b m/z | HPLC-Fraction | |
|---|---|---|---|---|---|
| 855.51 | 256.10 (C15) | 600.16 | 24 | ||
| 228.09 (C13) | 628.18 | ||||
| 926.55 | 256.0 | 671.42 | 24 | ||
| 869.53 | 256.15 (C15) | 614.12 | 25 | ||
| 228.18 (C13) | 642.21 | ||||
| 940.57 | 256.09 | 685.15 | 24 | ||
| A | 883.53 | 256.25 | 628.32 | 26 | |
| A | 954.57 | 256.21 | 699.21 | 25 | |
| B | 897.58 | 256.29 | 642.38 | 27 | |
| B | 968.62 | 256.19 | 713.22 | 25 | |
| E | 911.55 | 256.26 | 656.16 | 27/28 | |
| E | 982.62 | 256.26 | 727.32 | 26 | |
| F | 925.58 | 256.22 | 670.22 | 28/29 | |
| F | 996.65 | 256.20 | 741.34 | 27/28 | |
| C | 947.51 | 256.28 | 692.28 | 25 | |
| C | 1018.58 | 256.21 | 763.19 | 25 | |
| D | 961.55 | 256.20 | 706.23 | 25 | |
| D | 1032.61 | 256.17 | 777.10 | 25 | |
| m/z + 16 | m/z + 71 | ||||
| A | 899.55 | 272.18 | 628.27 | 22 | |
| A | 970.51 | 272.13 | 699.22 | 22/23 | |
| B | 913.57 | 272.12 | 642.16 | 22/23 | |
| B | 984.52 | 272.15 | 713.25 | 22/23 | |
| m/z + 14 | |||||
| 897.25 | 270.15 | 628.25 | 23 | ||
| 911.24 | 270.10 | 642.16 | 23 |
| Strain DSM 32871: Polymyxin E | Strain M1: Polymyxin P | |
|---|---|---|
| [M + H, Na, K]+ | E1: 1155.7/1177.8/1193.8 | P1: 1177.4/1199.3/1215.4 |
| E2: 1169.8/1191.8/1207.8 | P2: 1191.4/1213.4/1229.3 | |
| [M − H + 2K]+ | E1: 1231.9 | P1: 1253.9 |
| E2: 1245.9 | P2: 1267.9 | |
| [M + H + 146]+ | E1: 1302.0 | P1: 1323.9 |
| E2: 1316.0 | P2: 1337.9 | |
| [M + H + 197]+ | E1: 1352.9 | |
| E2: 1366.9 |
| (a) Dipeptide Fragments | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| m/z | ||||||||||||
| calc. | found | |||||||||||
| C9-FA-Dab | 141.13 | 141.03 | ||||||||||
| Dab-Thr | 202.12 | 202.02 | ||||||||||
| Thr-Dab | 202.12 | 202.02 | ||||||||||
| Dab-Thr | 202.12 | 202.02 | ||||||||||
| Thr-Dab | 202.12 | 202.02 | ||||||||||
| Dab-Leu | 214.16 | 214.06 | ||||||||||
| Leu-Leu | 227.18 | 227.08 | ||||||||||
| Leu-Dab | 214.16 | 214.06 | ||||||||||
| Dab-Dab | 201.13 | 201.03 | ||||||||||
| Dab-Thr | 202.12 | 202.02 | ||||||||||
| Thr + 145.41 | 247.47 | − | ||||||||||
| (b) Tripeptide Fragments | ||||||||||||
| C9-FA-Dab-Thr | 342.24 | 324.18 * | ||||||||||
| Dab-Thr-Dab | 302.18 | 302.14 | ||||||||||
| Thr-Dab-Thr | 303.17 | 285.11 * | ||||||||||
| Dab-Thr-Dab | 302.18 | 302.14 | ||||||||||
| Thr-Dab-Leu | 315.20 | 297.17 * | ||||||||||
| Dab-Leu-Leu | 327.24 | 327.21 | ||||||||||
| Leu-Leu-Dab | 327.24 | 327.21 | ||||||||||
| Leu-Dab-Dab | 314.22 | 314.18 | ||||||||||
| Dab-Dab-Thr | 302.17 | 302.14 | ||||||||||
| Dab-Thr + 145.41 | 347.43 | − | ||||||||||
| (c) Tetrapeptide Fragments | ||||||||||||
| C9-FA-Dab-Thr-Dab | 442.30 | 442.20 | ||||||||||
| Dab-Thr-Dab-Thr | 403.23 | 403.23 | ||||||||||
| Thr-Dab-Thr-Dab | 403.23 | 403.23 | ||||||||||
| Dab-Thr-Dab-Leu | 415.27 | 415.25 | ||||||||||
| Thr-Dab_Leu-Leu | 428.29 | 410.28 * | ||||||||||
| Dab-Leu-Leu-Dab | 427.30 | 427.26 | ||||||||||
| Leu-Leu-Dab-Dab | 427.30 | 427.26 | ||||||||||
| Leu-Dab-Dab-Thr | 415.27 | 415.25 | ||||||||||
| Dab-Dab-Thr + 145.41 | 442.59 | 447.31 | ||||||||||
| (d) Pentapeptide Fragments | ||||||||||||
| C9-FA-Dab-Thr-Dab-Thr | 543.35 | 507.28 ** | ||||||||||
| Dab-Thr-Dab-Thr-Dab | 503.35 | 503.25 | ||||||||||
| Thr-Dab-Thr-Dab-Leu | 516.31 | 498.25 * | ||||||||||
| Dab-Thr-Dab-Leu-Leu | 528.35 | 528.35 | ||||||||||
| Thr-Dab-Leu-Leu-Dab | 528.35 | 528.35 | ||||||||||
| Dab-Leu-Leu-Dab-Dab | 527.37 | − | ||||||||||
| Leu-Leu.Dab-Dab-Thr | 528.35 | 528.35 | ||||||||||
| Leu-Dab-Dab-Thr + 145.41 | 560.68 | − | ||||||||||
| M1 | DSM32871 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Kovats Index | NA | LB | TSA | LANDY | NA | LB | TSA | LANDY | Biological Function | Reference |
| Methanethiol | 400 | x | x | x | Broccoli and lettuce seed germination inhibition | Lo Cantore et al., 2015 [70] | |||||
| Acetone | 455 | x | x | x | x | x | Potential contribution to inhibition of diverse fungi | Stotzky and Schenck, 1976 [71] | |||
| 1,4-Pentadiene | 518 | x | x | x | x | x | n.a. | ||||
| n-Hexane | 602 | x | n.a. | ||||||||
| 2-Methyl-1-propanol * | 618 | x | n.a. | ||||||||
| 3-Propoxy-1-propene * | 646 | x | x | n.a. | |||||||
| 3-Methylpentane | 650 | x | x | A biomarker for a breath test for detection of caries | Hertel et al., 2016 [72] | ||||||
| 2-Methylbutanal | 656 | x | x | x | n.a. | ||||||
| 2-Methylbutanenitrile * | 699 | x | n.a. | ||||||||
| Acetoin | 709 | x | x | x | x | x | x | x | Plant growth promotion and ISR activation in A. thaliana, increase of root length and number of lateral roots in Lactuca sativa | Ryu et al., 2003, 2004; Rudrappa et al., 2010; Fincheira et al., 2016 [50,54,65,68] | |
| 1-Chloropentane | 725 | x | n.a. | ||||||||
| 2-Methyl-1-butanol | 726 | x | x | x | x | x | x | Inhibtion of appressoria germination and germination of Phyllosticta citricarpa | Toffano et al., 2017 [64] | ||
| 3-Methyl-1-butanol | 727 | x | x | x | x | x | x | Growth promotion and enhanced salinity tolerance of Arabidopsis thaliana | Ledger et al., 2016 [67] | ||
| Dimethyl sulfone | 733 | x | n.a. | ||||||||
| 2,3-Butanediol | 743 | x | x | Enhanced growth of tobacco plants and ISR against Erwinia carotovora | Han et al., 2006 [66] | ||||||
| Butyl acetate | 802 | x | n.a. | ||||||||
| 3-Methylbutyl acetate | 864 | x | n.a. | ||||||||
| 2-Heptanone | 881 | x | n.a. | ||||||||
| 1-Octen-3-ol | 969 | x | x | Inhibit the development of Arabidopsis thaliana and modify its oxidative metabolism | Splivallo et al., 2007 [73] | ||||||
| 2-Ethyl-1-hexanol | 1018 | x | x | x | Inhibition of mycelial growth of S. sclerotiorum and reduction of sclerotial germination | Fernando et al., 2005 [62] | |||||
| 2-Isopropyl-3-methylpyrazine * | 1033 | x | x | x | x | x | n.a. | ||||
| 2-Isopropyl-5-methylpyrazine | 1034 | x | n.a. | ||||||||
| 3-Isopropyl-2,5-dimethylpyrazine | 1129 | x | x | x | n.a. | ||||||
| 2-Decanone | 1152 | x | x | x | x | x | x | Inhibition of mycelial growth of Rhizoctonia solani | Weise et al., 2012 [74] | ||
| 2-Decanol | 1166 | x | x | x | x | n.a. | |||||
| 2,3-Diethyl-5-methylpyrazine * | 1261 | x | x | x | n.a. | ||||||
| 2-Isobutyl-3-isopropylpyrazine * | 1267 | x | x | x | x | x | x | n.a. | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mülner, P.; Schwarz, E.; Dietel, K.; Herfort, S.; Jähne, J.; Lasch, P.; Cernava, T.; Berg, G.; Vater, J. Fusaricidins, Polymyxins and Volatiles Produced by Paenibacillus polymyxa Strains DSM 32871 and M1. Pathogens 2021, 10, 1485. https://doi.org/10.3390/pathogens10111485
Mülner P, Schwarz E, Dietel K, Herfort S, Jähne J, Lasch P, Cernava T, Berg G, Vater J. Fusaricidins, Polymyxins and Volatiles Produced by Paenibacillus polymyxa Strains DSM 32871 and M1. Pathogens. 2021; 10(11):1485. https://doi.org/10.3390/pathogens10111485
Chicago/Turabian StyleMülner, Pascal, Elisa Schwarz, Kristin Dietel, Stefanie Herfort, Jennifer Jähne, Peter Lasch, Tomislav Cernava, Gabriele Berg, and Joachim Vater. 2021. "Fusaricidins, Polymyxins and Volatiles Produced by Paenibacillus polymyxa Strains DSM 32871 and M1" Pathogens 10, no. 11: 1485. https://doi.org/10.3390/pathogens10111485