
Impact of Fluid Flow Shear Stress on Osteoblast Differentiation and Cross-Talk with Articular Chondrocytes

 and
and
Abstract
:1. Introduction
2. Results
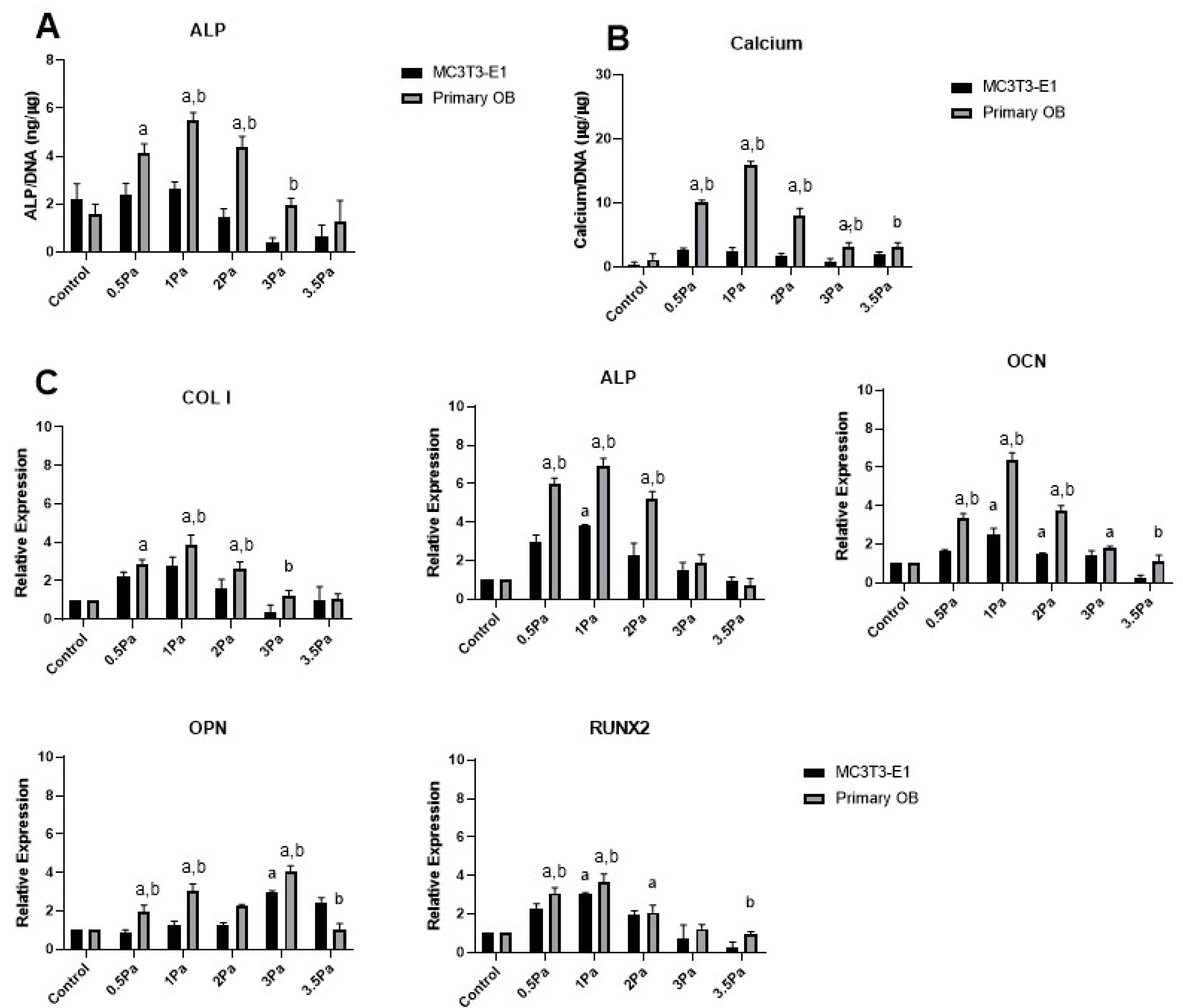
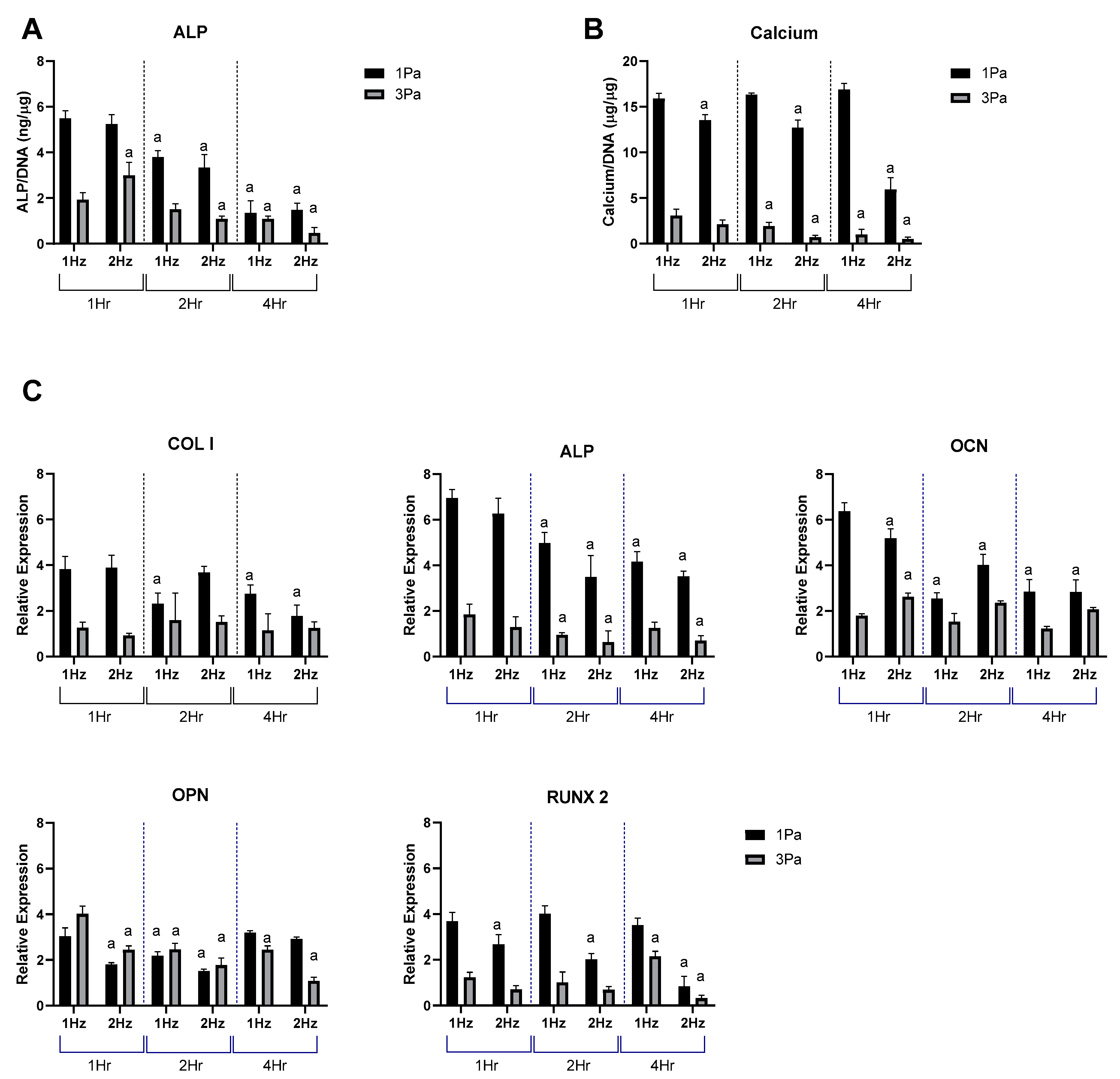
2.1. Effects of Fluid Flow Shear Stress (FFSS) on the Proliferation and Differentiation of Primary and Cell Line Osteoblasts
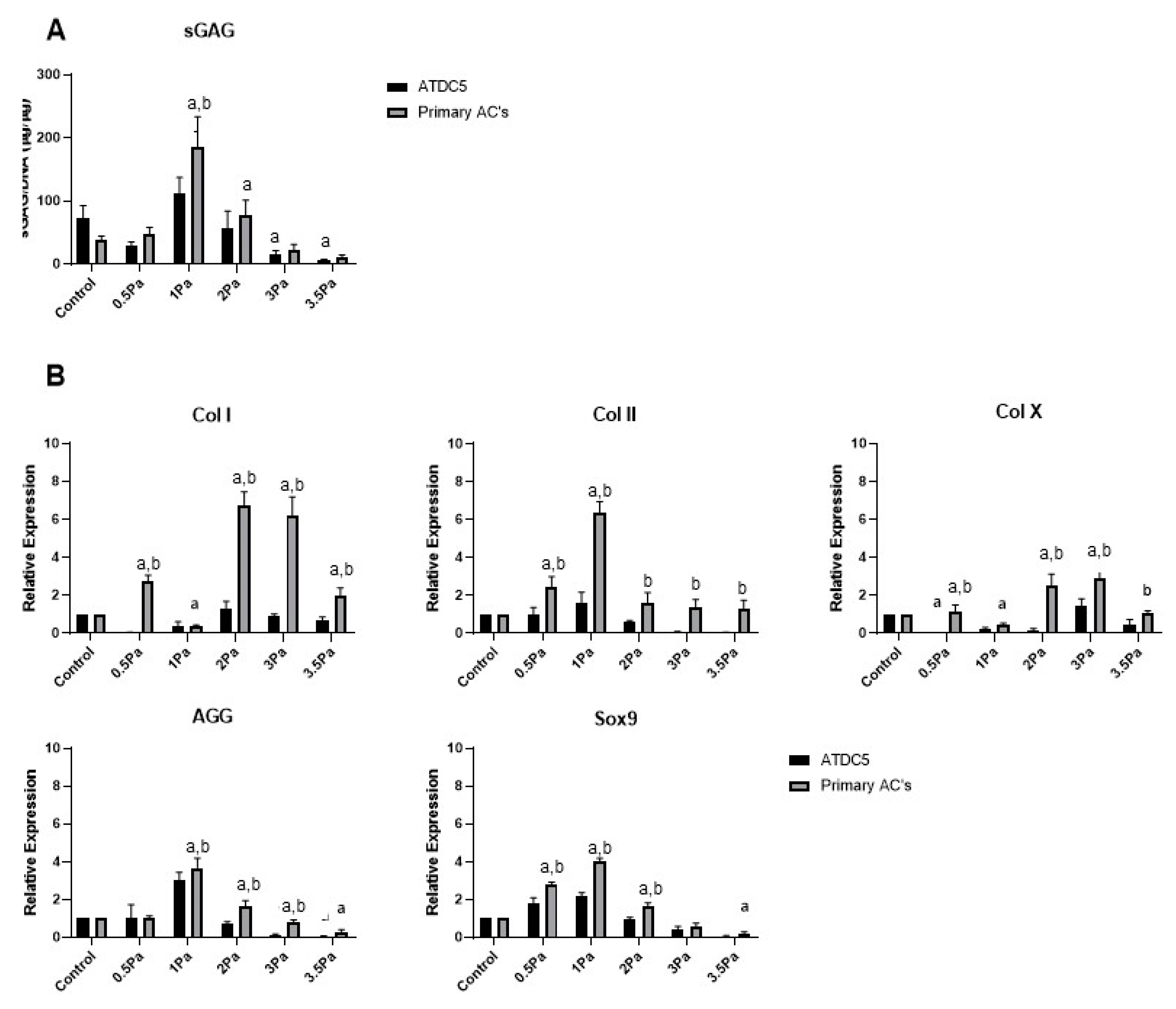
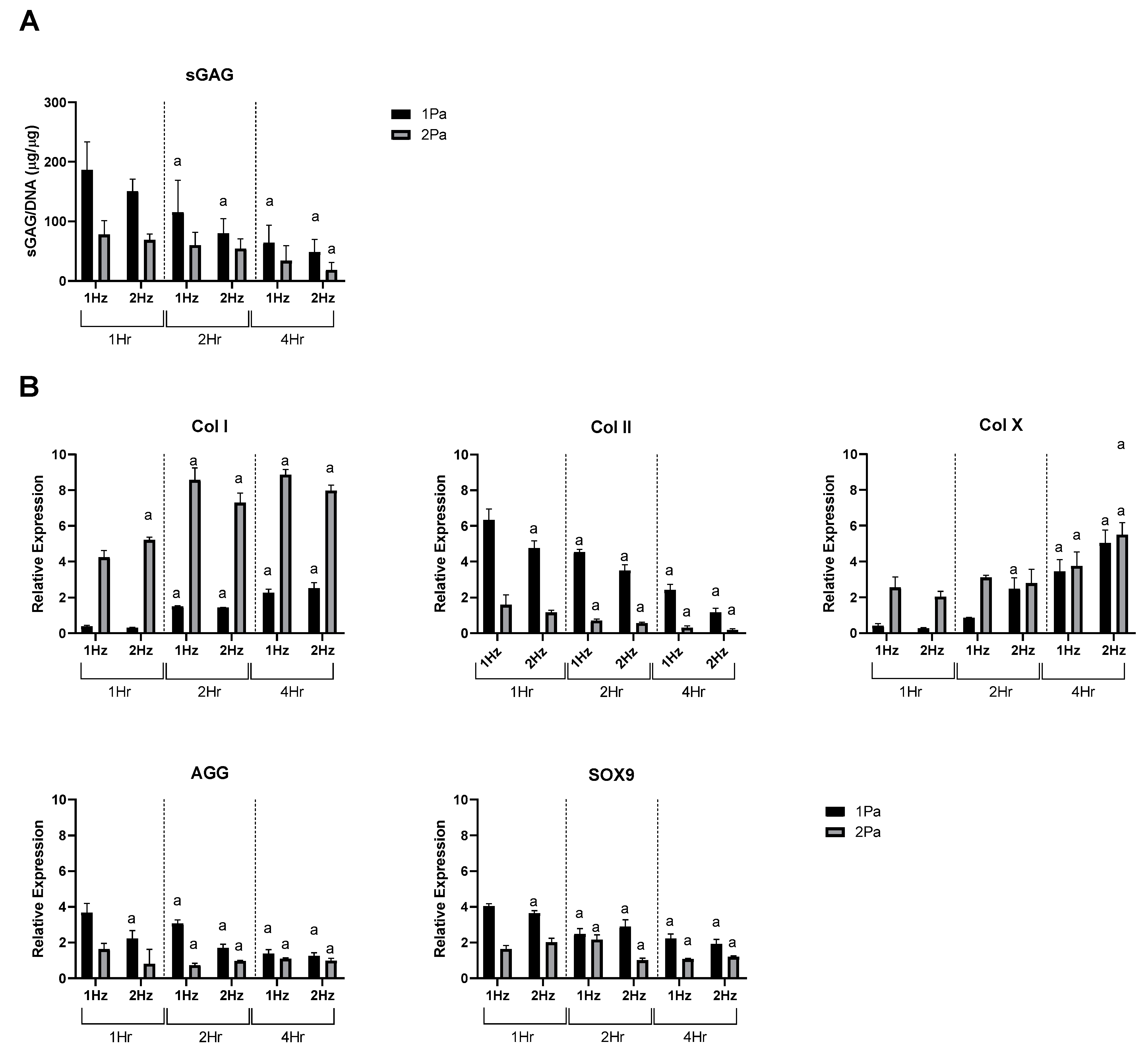
2.2. Effects of FFSS on the Proliferation and Differentiation of Chondrocytes
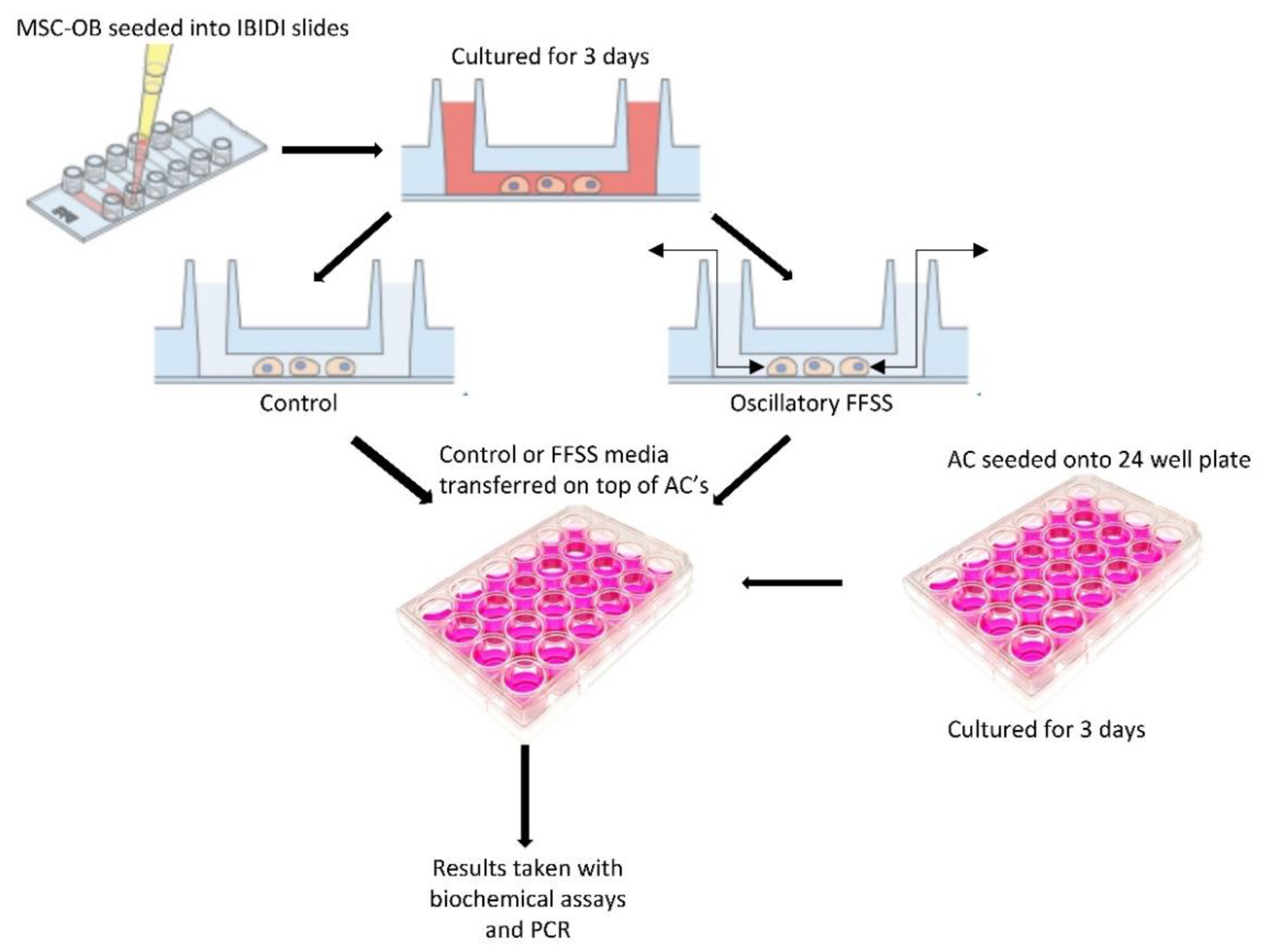
2.3. Effect of FFSS Induced Osteoblast Derived Soluble Factors on Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.1.1. Cell Line Culture
4.1.2. Primary Osteoblast Isolation
4.1.3. Primary Chondrocyte Isolation
4.2. In Vitro Fluid Flow Shear Stress Model
4.2.1. Preparation of Conditioned Medium (CM) from Osteoblasts
4.2.2. Experimental Set-Up for Conditioned Media Studies
4.3. Biochemical Assays
4.3.1. Biochemical Analysis of Osteoblast Cells
4.3.2. Biochemical Analysis of Chondrocyte Cells
4.4. qPCR
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenkre, J.S.; Bassett, J.H.D. The bone remodelling cycle. Ann. Clin. Biochem. 2018, 55, 308–327. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
- Dallas, S.L.; Bonewald, L.F. Dynamics of the transition from osteoblast to osteocyte. Ann. N. Y. Acad. Sci. 2010, 1192, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Bellido, T.; Plotkin, L.I.; Bruzzaniti, A. Bone Cells. In Basic and Applied Bone Biology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 37–55. [Google Scholar]
- Hinton, P.V.; Rackard, S.M.; Kennedy, O.D. In Vivo Osteocyte Mechanotransduction: Recent Developments and Future Directions. Curr. Osteoporos. Rep. 2018, 16, 746–753. [Google Scholar] [CrossRef]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef]
- Maes, C.; Kronenberg, H. Chapter 60—Bone Development and Remodeling. In Endocrinology: Adult and Pediatric, 7th ed.; Saunders: Philadelphia, PA, USA, 2016; pp. 1038–1062.e8. [Google Scholar]
- Ramme, A.J.; Lendhey, M.; Raya, J.G.; Kirsch, T.; Kennedy, O.D. A novel rat model for subchondral microdamage in acute knee injury: A potential mechanism in post-traumatic osteoarthritis. Osteoarthr. Cartil. 2016, 24, 1776–1785. [Google Scholar] [CrossRef] [Green Version]
- Selvarajah, L.; Curtis, A.M.; Kennedy, O.D. Bone Microdamage in Acute Knee Injury. Curr. Rheumatol. Rep. 2018, 20, 89. [Google Scholar] [CrossRef]
- Coughlin, T.R.; Kennedy, O.D. The role of subchondral bone damage in post-traumatic osteoarthritis. Ann. N. Y. Acad. Sci. 2016, 1383, 58–66. [Google Scholar] [CrossRef]
- Burr, D.B.; Radin, E.L. Microfractures and microcracks in subchondral bone: Are they relevant to osteoarthrosis? Rheum. Dis. Clin. N. Am. 2003, 29, 675–685. [Google Scholar] [CrossRef]
- Wang, X.; Masse, D.B.; Leng, H.; Roeder, R.K.; Niebur, G.L. Detection Of Trabecular Bone Microdamage by Micro-CT Imaging. J. Biomech. 2007, 40, 3397–3403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinbaum, S.; Cowin, S.C.; Zeng, Y. A model for the excitation of osteocytes by mechanical loading-induced bone fluid shear stresses. J. Biomech. 1994, 27, 339–360. [Google Scholar] [CrossRef]
- Klein-Nulend, J.; Bakker, A.D.; Bacabac, R.G.; Vatsa, A.; Weinbaum, S. Mechanosensation and transduction in osteocytes. Bone 2013, 54, 182–190. [Google Scholar] [CrossRef]
- Eschweiler, J.; Horn, N.; Rath, B.; Betsch, M.; Baroncini, A.; Tingart, M.; Migliorini, F. The Biomechanics of Cartilage—An Overview. Life 2021, 11, 302. [Google Scholar] [CrossRef]
- Pavalko, F.M.; Chen, N.X.; Turner, C.H.; Burr, D.B.; Atkinson, S.; Hsieh, Y.F.; Qiu, J.; Duncan, R.L. Fluid shear-induced mechanical signaling in MC3T3-E1 osteoblasts requires cytoskeleton-integrin interactions. Am. J. Physiol. 1998, 275, 1591–1601. [Google Scholar] [CrossRef]
- You, J.; Reilly, G.C.; Zhen, X.; Yellowley, C.E.; Chen, Q.; Donahue, H.J.; Jacobs, C.R. Osteopontin Gene Regulation by Oscillatory Fluid Flow via Intracellular Calcium Mobilization and Activation of Mitogen-activated Protein Kinase in MC3T3-E1 Osteoblasts. J. Biol. Chem. 2001, 276, 13365–13371. [Google Scholar] [CrossRef] [Green Version]
- Morinobu, M.; Ishijima, M.; Rittling, S.R.; Tsuji, K.; Yamamoto, H.; Nifuji, A.; Denhardt, D.T.; Noda, M. Osteopontin expression in osteoblasts and osteocytes during bone formation under mechanical stress in the calvarial suture in vivo. J. Bone Miner. Res. 2003, 18, 1706–1715. [Google Scholar] [CrossRef]
- Bakker, A.D.; Klein-Nulend, J. Osteoblast isolation from murine calvaria and long bones. Methods Mol. Biol. 2012, 816, 19–29. [Google Scholar]
- Manolagas, S.C. Osteocalcin promotes bone mineralization but is not a hormone. PLoS Genet. 2020, 16, e1008714. [Google Scholar] [CrossRef]
- Czekanska, E.M.; Stoddart, M.J.; Ralphs, J.R.; Richards, R.G.; Hayes, J.S. A phenotypic comparison of osteoblast cell lines versus human primary osteoblasts for biomaterials testing. J. Biomed. Mater. Res. Part A 2014, 102, 2636–2643. [Google Scholar] [CrossRef]
- Takeuchi, K.; Abe, M.; Hiasa, M.; Oda, A.; Amou, H.; Kido, S.; Harada, T.; Tanaka, O.; Miki, H.; Nakamura, S.; et al. TGF-β inhibition restores terminal osteoblast differentiation to suppress myeloma growth. PLoS ONE 2010, 5, e9870. [Google Scholar] [CrossRef]
- Sugawara, Y.; Suzuki, K.; Koshikawa, M.; Ando, M.; Iida, J. Necessity of enzymatic activity of alkaline phosphatase for mineralization of osteoblastic cells. Jpn. J. Pharmacol. 2002, 88, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Orimo, H. The Mechanism of Mineralization and the Role of Alkaline Phosphatase in Health and Disease. J. Nippon Med. Sch. 2010, 77, 42–43. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Kumar, C.; Bohl, S.; Klingmueller, U.; Mann, M. Comparative Proteomic Phenotyping of Cell Lines and Primary Cells to Assess Preservation of Cell Type-specific Functions. Mol. Cell. Proteom. 2009, 8, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Salvat, C.; Pigenet, A.; Humbert, L.; Berenbaum, F.; Thrion, S. Immature murine articular chondrocytes in primary culture: A new tool for investigating cartilage. Osteoarthr. Cartil. 2005, 13, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Gosset, M.; Berenbaum, F.; Thirion, S.; Jacques, C. Primary culture and phenotyping of murine chondrocytes. Nat. Protoc. 2008, 3, 1253–1260. [Google Scholar] [CrossRef]
- Hoffman, B.E.; Newman-Tarr, T.M.; Gibbard, A.; Wang, S.; Hanning, C.; Pratta, M.A.; Boyle, R.J.; Kumar, S.; Majumdar, M.K. Development and characterization of a human articular cartilage-derived chondrocyte cell line that retains chondrocyte phenotype. J. Cell. Physiol. 2010, 222, 695–702. [Google Scholar] [CrossRef]
- Kartsogiannis, V.; Ng, K.W. Cell lines and primary cell cultures in the study of bone cell biology. Mol. Cell. Endocrinol. 2004, 228, 79–102. [Google Scholar] [CrossRef]
- Xing, J.; Li, Y.; Lin, M.; Wang, J.; Wu, J.; Ma, Y.; Wang, Y.; Yang, L.; Luo, Y. Surface chemistry modulates osteoblasts sensitivity to low fluid shear stress. J. Biomed. Mater. Res. Part A 2014, 102, 4151–4160. [Google Scholar] [CrossRef]
- Ban, Y.; Wu, Y.-Y.; Yu, T.; Geng, N.; Wang, Y.-Y.; Liu, X.-G.; Gong, P. Response of osteoblasts to low fluid shear stress is time dependent. Tissue Cell 2011, 43, 311–317. [Google Scholar] [CrossRef]
- Wittkowske, C.; Reilly, G.C.; Lacroix, D.; Perrault, C.M. In Vitro Bone Cell Models: Impact of Fluid Shear Stress on Bone Formation. Front. Bioeng. Biotechnol. 2016, 4, 87. [Google Scholar] [CrossRef] [Green Version]
- Liegibel, U.M.; Sommer, U.; Bundschuh, B.; Schweizer, B.; Hilscher, U.; Lieder, A.; Nawroth, P.; Kasperk, C. Fluid shear of low magnitude increases growth and expression of TGFβ1 and adhesion molecules in human bone cells in vitro. Exp. Clin. Endocrinol. Diabetes 2004, 112, 356–363. [Google Scholar] [CrossRef]
- Owan, I.; Burr, D.B.; Turner, C.H.; Qiu, J.; Tu, Y.; Onyia, J.E.; Duncan, R.L. Mechanotransduction in bone: Osteoblasts are more responsive to fluid forces than mechanical strain. Am. J. Physiol.-Cell Physiol. 1997, 273, C810–C815. [Google Scholar] [CrossRef]
- Genetos, D.C.; Geist, D.J.; Liu, D.; Donahue, H.J.; Duncan, R.L. Fluid shear-induced ATP secretion mediates prostaglandin release in MC3T3-E1 osteoblasts. J. Bone Miner. Res. 2005, 20, 41–49. [Google Scholar] [CrossRef]
- Thi, M.M.; Islam, S.; Suadicani, S.O.; Spray, D.C. Connexin43 and Pannexin1 Channels in Osteoblasts: Who is the “hemichannel”? J. Membr. Biol. 2012, 245, 401. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Rose, E.; Frances, D.; Sun, Y.; You, L. Effect of oscillating fluid flow stimulation on osteocyte mRNA expression. J. Biomech. 2012, 45, 247–251. [Google Scholar] [CrossRef]
- Gemmiti, C.V.; Guldberg, R.E. Fluid flow increases type II collagen deposition and tensile mechanical properties in bioreactor-grown tissue-engineered cartilage. Tissue Eng. 2006, 12, 469–479. [Google Scholar] [CrossRef]
- Liao, J.; Guo, X.; Nelson, D.; Kasper, F.K.; Mikos, A.G. Modulation of osteogenic properties of biodegradable polymer/extracellular matrix scaffolds generated with a flow perfusion bioreactor. Acta Biomater. 2010, 6, 2386–2393. [Google Scholar] [CrossRef] [Green Version]
- Kock, L.M.; Malda, J.; Dhert, W.J.A.; Ito, K.; Gawlitta, D. Flow-perfusion interferes with chondrogenic and hypertrophic matrix production by mesenchymal stem cells. J. Biomech. 2014, 47, 2122–2129. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, N.; Gharravi, A.M. Shear bioreactors stimulating chondrocyte regeneration, a systematic review. Inflamm. Regen. 2019, 39, 16. [Google Scholar] [CrossRef] [Green Version]
- Gharravi, A.M.; Orazizadeh, M.; Hashemitabar, M. Fluid-induced low shear stress improves cartilage like tissue fabrication by encapsulating chondrocytes. Cell Tissue Bank. 2016, 17, 117–122. [Google Scholar] [CrossRef]
- Carmona-Moran, C.A.; Wick, T.M. Transient Growth Factor Stimulation Improves Chondrogenesis in Static Culture and Under Dynamic Conditions in a Novel Shear and Perfusion Bioreactor. Cell. Mol. Bioeng. 2015, 8, 267–277. [Google Scholar] [CrossRef]
- Dahlin, R.L.; Meretoja, V.V.; Ni, M.; Kasper, F.K.; Mikos, A.G. Hypoxia and flow perfusion modulate proliferation and gene expression of articular chondrocytes on porous scaffolds. AIChE J. 2013, 59, 3158–3166. [Google Scholar] [CrossRef]
- Zhu, F.; Wang, P.; Lee, N.H.; Goldring, M.B.; Konstantopoulos, K. Prolonged Application of High Fluid Shear to Chondrocytes Recapitulates Gene Expression Profiles Associated with Osteoarthritis. PLoS ONE 2010, 5, e15174. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Zhou, X.; Li, W.; Novotny, J.E.; Doty, S.B.; Wang, L. In situ measurement of transport between subchondral bone and articular cartilage. J. Orthop. Res. 2009, 27, 1347–1352. [Google Scholar] [CrossRef] [Green Version]
- Arkill, K.P.; Winlove, C.P. Solute transport in the deep and calcified zones of articular cartilage. Osteoarthr. Cartil. 2008, 16, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Carpintero-Fernandez, P.; Gago-Fuentes, R.; Wang, H.Z.; Fonseca, E.; Caeiro, J.R.; Valiunas, V.; Brink, P.R.; Mayan, M.D. Intercellular communication via gap junction channels between chondrocytes and bone cells. Biochim. Biophys. Acta—Biomembr. 2018, 1860, 2499–2505. [Google Scholar] [CrossRef]
- µ-Slide VI 0.4|Channel Slide for Parallel Flow Assays|Ibidi. Available online: https://ibidi.com/channel-slides/57--slide-vi-04.html (accessed on 10 August 2021).
- Demitri, C.; Raucci, M.G.; Giuri, A.; De Benedictis, V.M.; Giugliano, D.; Calcagnile, P.; Sannino, A.; Ambrosio, L. Cellulose-based porous scaffold for bone tissue engineering applications: Assessment of hMSC proliferation and differentiation. J. Biomed. Mater. Res. Part A 2016, 104, 726–733. [Google Scholar] [CrossRef]
- Wang, D.; Christensen, K.; Chawla, K.; Xiao, G.; Krebsbach, P.H.; Franceschi, R.T. Isolation and characterization of MC3T3-E1 preosteoblast subclones with distinct in vitro and in vivo differentiation/mineralization potential. J. Bone Miner. Res. 1999, 14, 893–903. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| τ [Pa] | Φ [mL/min] | Displacement [µL] |
|---|---|---|
| 0.5 | 3.94 | 63 |
| 1 | 7.89 | 127 |
| 2 | 15.77 | 253 |
| 3 | 23.66 | 380 |
| 3.5 | 27.60 | 447 |
| Primer Name | Forward Sequence | Reverse Sequence | Accession Number |
|---|---|---|---|
| Mouse Collagen I | 5′ CTTCACCTACAGCACCCTTGTG 3′ | 5′ GATGACTGTCTTGCCCCAAGTT 3′ | NM_007742.4 |
| Mouse Collagen II | 5′AAGTCACTGAACAACCGATTGAGA 3′ | 5′ AATGCGAGCAGGGTTCTTG 3′ | NM_031163.4 |
| Mouse Collagen X | 5′ TTCTCCTACCACGTGCATGTG 3′ | 5′ AGGCGTGCCGTTCTTATACAG 3′ | BC_15156930.1 |
| Mouse Aggrecan | 5′ GCCACGGTGCCCTTTTTAC 3′ | 5′ GAGAGAGGCGAATGGAACGA 3′ | NM_007424.34 |
| Mouse SOX9 | 5′ GGTGGAGTAGAGCCCTGAGC 3′ | 5′ CCTTCAACCTTCCTCACTACAGC 3′ | NM_011448.4 |
| Mouse OCN | 5′ AAGCAGGAGGGCAATAAGGT 3′ | 5 ′ TTTGTAGGCGGTCTTCAAGC 3′ | L24429.1 |
| Mouse OPN | 5′ AGCAAGAAACTCTTCCAAGCAA 3′ | 5′ GTGAGATTCGTCAGATTCATCCG 3′ | J04806.1 |
| Mouse OPN | 5′ AGCAAGAAACTCTTCCAAGCAA 3′ | 5′ GTGAGATTCGTCAGATTCATCCG 3′ | NM_0012040201.1 |
| Mouse RUNX2 | 5′ ACTCTTCTGGAG CCGTTTATG 3′ | 5′ GTGAATCTGGCC ATGTTTGTG 3′ | NM_001146038.2 |
| Mouse ALP | 5′ AACCCAGACAC AAGCATTCC 3′ | 5′ GAGAGCGAAGGG TCAGTCAG 3′ | NM_007431.4 |
| Mouse BMP2 | 5′ ACACAGCTGGTCACAGATAAG 3′ | 5′ CTTCCGCTGTTTGTGTTTGG 3′ | L25602.1 |
| Mouse 18S | 5′ ACGAGACTCTGGCATGCTAACTAGT 3′ | 5′ CGCCACTTGTCCCTCTAAGAA 3′ | 7SYS_2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinton, P.V.; Genoud, K.J.; Early, J.O.; O’Brien, F.J.; Kennedy, O.D. Impact of Fluid Flow Shear Stress on Osteoblast Differentiation and Cross-Talk with Articular Chondrocytes. Int. J. Mol. Sci. 2022, 23, 9505. https://doi.org/10.3390/ijms23169505
Hinton PV, Genoud KJ, Early JO, O’Brien FJ, Kennedy OD. Impact of Fluid Flow Shear Stress on Osteoblast Differentiation and Cross-Talk with Articular Chondrocytes. International Journal of Molecular Sciences. 2022; 23(16):9505. https://doi.org/10.3390/ijms23169505
Chicago/Turabian StyleHinton, Paige V., Katelyn J. Genoud, James O. Early, Fergal J. O’Brien, and Oran D. Kennedy. 2022. "Impact of Fluid Flow Shear Stress on Osteoblast Differentiation and Cross-Talk with Articular Chondrocytes" International Journal of Molecular Sciences 23, no. 16: 9505. https://doi.org/10.3390/ijms23169505