Capture and Release of Phosphorus by Periphyton in Closed Water Systems Influenced by Illumination and Temperature

1
State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
2
Hubei Academy of Environmental Sciences, Wuhan 430072, China
3
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Water 2019, 11(5), 1021; https://doi.org/10.3390/w11051021
Submission received: 8 April 2019
/
Revised: 27 April 2019
/
Accepted: 14 May 2019
/
Published: 16 May 2019
(This article belongs to the Section Water Quality and Contamination)
Abstract
:Periphyton is known to play an important role in the self-purification of aquatic ecosystems. However, little attention has been paid to the understanding of P distribution and its partitioning influenced by the physical parameters when periphyton is separated from the sediment. In this work, the effect of periphyton on the capture and release of phosphorus in closed water systems was studied and the influence of illumination and temperature conditions were investigated. Results showed that phosphorus was transferred from water to periphyton during the experiment at 15 °C, but periphyton turned from a sink to a source of phosphorus in a few days at 25 and 35 °C. Phosphorus capture in periphyton was more enhanced when illuminated at 70 than 20 μmol photons m−2 s−1 at 25 and 35 °C, but not at 15 °C. At the end of the experiment, cyanobacteria became more abundant at 25 and 35 °C and phosphorus fractionation showed that labile-P was predominant in periphyton. The release of the captured phosphorus could be related to the disaggregation of periphyton following the depletion of nutrients. Therefore, periphyton act as a temporary storage of phosphorus following nutrient input in closed water systems and the capture and release of phosphorus is strongly influenced by the environmental conditions.
1. Introduction
Closed water systems, such as ponds, paddies, and lagoons, are sensitive to nutrient input. Nutrients from point or non-point sources can lead to a significant change in water quality in these closed water systems. Microorganisms such as algae and bacteria can response quickly to such changes due to their rapid propagation. Periphyton, with microbial assemblages consisting of algae, bacteria, fungi, protozoa, metazoan, and detritus, can develop rapidly in these closed water systems under suitable environmental conditions [1,2]. Periphyton is considered to be an important sink for phosphorus (P) and participates actively in P retention and turnover. The diverse microbial community in periphyton also plays a significant role in the self-purification of aquatic ecosystems [3,4,5]. Many technologies are currently in place to remove excess P from waters, including biological methods using periphyton as a subject of great interest [6,7,8].
Development and function of periphyton have been found to be affected by many factors such as nutrient levels, environmental conditions, and grazing [9,10,11] and these factors can have interactive effects on periphyton [12,13,14]. Generally, higher nutrient level, suitable illumination, moderate temperature, and lower disturbance favor the development of periphyton while lower nutrients, grazing pressure, higher disturbance, and extreme (too high or too low) illumination or temperature conditions can inhibit periphyton growth. Among these factors, illumination and temperature are important and more easily controlled variables for affecting the P cycle, but their interactive effects on periphytons are complex and are not well characterized [15].
In closed waters, nutrient loadings are retained in the system and are subjected to physicochemical and biological transformation processes. Nitrogen can leave the system via denitrification, while phosphorus can only change between different forms. Phosphorus capture and release in periphyton could change under the influence of illumination and temperature. Previous research mainly focused on open water systems; few studies have investigated the role of periphyton in phosphorus transformation in closed water systems. Therefore, the purpose of this study is to characterize the temporal changes of periphyton in a closed water system following nutrient enrichment and to investigate the capacity of periphyton to capture P from overlying water and the potential for P to be released back to the water column. The influence of two important environmental factors (illuminance and temperature) and their interactive effects on periphyton and the subsequent effects on the P cycle were studied.
2. Materials and Methods
2.1. Experimental Setup
Three water baths (120 × 90 × 30 cm) were used for the experiment. The temperature of each water bath was maintained using a circulating water temperature control system and were 15, 25, and 35 °C, respectively. Each microcosm was illuminated with light-emitting diodes (LED) lights with a 16 to 8 h light–dark cycle. Each water bath was divided into two parts and the photosynthetically active radiation (PAR) of each part at the water surface was 20 and 70 μmol photons m−2 s−1, respectively. This experimental setup resulted in six treatments, including H15 (70 μmol photons m−2 s−1, 15 °C), H25 (70 μmol photons m−2 s−1, 25 °C), H35 (70 μmol photons m−2 s−1, 35 °C), L15 (20 μmol photons m−2 s−1, 15 °C), L25 (20 μmol photons m−2 s−1, 25 °C), and L35 (20 μmol photons m−2 s−1, 35 °C).
A total of 108 plastic boxes (150 × 105 × 94 mm) were used as microcosms for closed water systems and 5 glass slides (76 × 25 × 2 mm) were placed on the bottom of each box as substrates for periphyton. Each box was filled with WC medium (https://utex.org/products/wc-medium) to sustain periphyton growth and inoculated with 10 mL slurry of periphyton collected from East Lake in May, which is a mesotrophic urban lake in Wuhan, China. The slurry was expanded in the laboratory at 25 °C, 20 ± 5 μmol photons m−2 s−1 as the temperature and light in East Lake, respectively. The slurry passed through a 0.064 mm mesh size net to remove large debris and zooplankton. The biomass of the slurry was 0.018 g mL−1. The periphyton was allowed to develop for 5 days at 25 °C, 20 ± 5 μmol photons m−2 s−1. Then the boxes for each treatment were moved to the corresponding water bath to start the experiment and the nutrient concentration was measured as the initial concentration.
2.2. Sampling and Analysis
The experiment lasted for 12 days and three plastic boxes were retrieved from each water bath on day 0, 1, 2, 4, 6, 9, and 12. In order to keep the same amount of water, deionized water was added to the boxes periodically to compensate for the evaporation loss.
In the water samples, dissolved oxygen (DO) and pH were measured using an HQ40d meter (Hach, Loveland, CO, USA) at the same time (9:00 am) on the sampling day. Total phosphorus (TP), total dissolved phosphorus (TDP), dissolved inorganic phosphorus (DIP), total nitrogen (TN), and NH4+-N of the water were determined following standard methods [16]. Particular phosphorus (PP) and dissolved organic phosphorus (DOP) were calculated by subtracting TDP from TP and by subtracting DIP from TDP, respectively. Initial concentrations of TP, TSP, DIP, DOP, PP, TN, and NH4+-N were measured and found to be 1.1, 0.93, 0.89, 0.09, 0.12, 7.34, and 4.18 mg/L, respectively.
Periphyton samples were removed from the glass slides with a spatula. Ash-free dry weight (AFDW) was determined gravimetrically after drying at 105 °C and burning at 450 °C. Chlorophyll a (Chl.a) was measured colorimetrically after extraction with 90% acetone. Alkaline phosphatase (APase) activity was measured following [17]. The content of TP in periphyton samples was determined after extraction with concentrated HCl [18]. Phosphorus was fractionated into loosely sorbed P (Labile-P), reductant-soluble P (BD-P), metal oxide-bound P (Fe/Al-P), calcium-bound P (Ca-P), and residual P (Res-P). Labile-P, BD-P, Fe/Al-P, and Ca-P were determined following sequential extraction with 1.0 M MgCl2, BD reagent (0.11 M dithionite buffered by bicarbonate), 0.1 M NaOH, and 0.5 M HCl and Res-P was calculated by subtracting all extractable P fractions from TP. On day 12, the algae composition of the periphyton samples was identified by referring to Hu and Wei [19].
2.3. Statistical Analysis
Differences among treatments and interactive effects between illumination, temperature, and time were analyzed using repeated measurement analysis of variance (ANOVA). Probabilities within groups (day and interactions) were corrected for sphericity using the Greenhouse–Geisser correction. All probabilities were adjusted by an LSD post-hoc test. Statistical analysis was performed using SPSS software Version 18.0 (SPSS Inc, Chicago, IL, USA). A confidence level of 95% was used. All figures were plotted by the GraphPad Prism 6 software. The results are expressed as mean ± standard deviation (SD) of three replicates and SD is indicated using error bars in the figures.
3. Results
3.1. Changes in DO and pH in Water
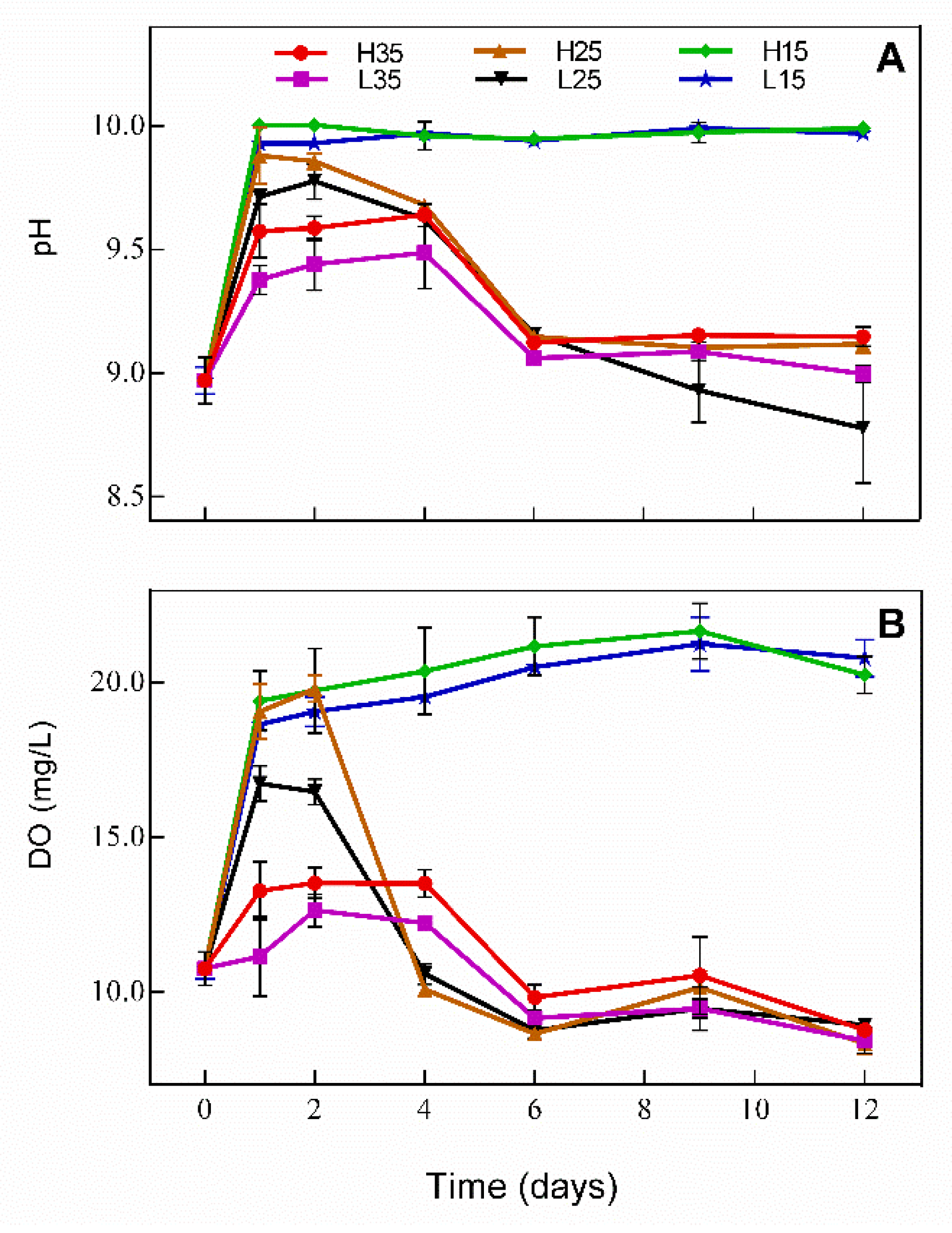
Changes in DO and pH in water during the experiment are presented in Figure 1 and follow similar trends. In H15 and L15, DO and pH increased significantly from day 0 to day 1 (p < 0.05) and were maintained at about 20 mg/L and 10 mg/L, respectively. For the rest of treatments, DO and pH increased first and then decreased. In H25 and L25, DO and pH began to drop on day 2, while in H35 and L35 DO and pH begun to drop on day 4; both changes were more marked in H25 and L25. Among treatments, DO and pH showed no statistically significant difference during the experiment between H15 and L15 (p > 0.05), and both were significantly higher in H25 than in L25 on day 1 and 2 (p < 0.05) and were higher in H35 than in L35 on day 1, 2, and 4 (p < 0.05). On day 12, no significant difference was observed among H35, L35, H25, and L25 for DO (p > 0.05), while L25 had the lowest pH.
3.2. Changes in Phosphorus in Water
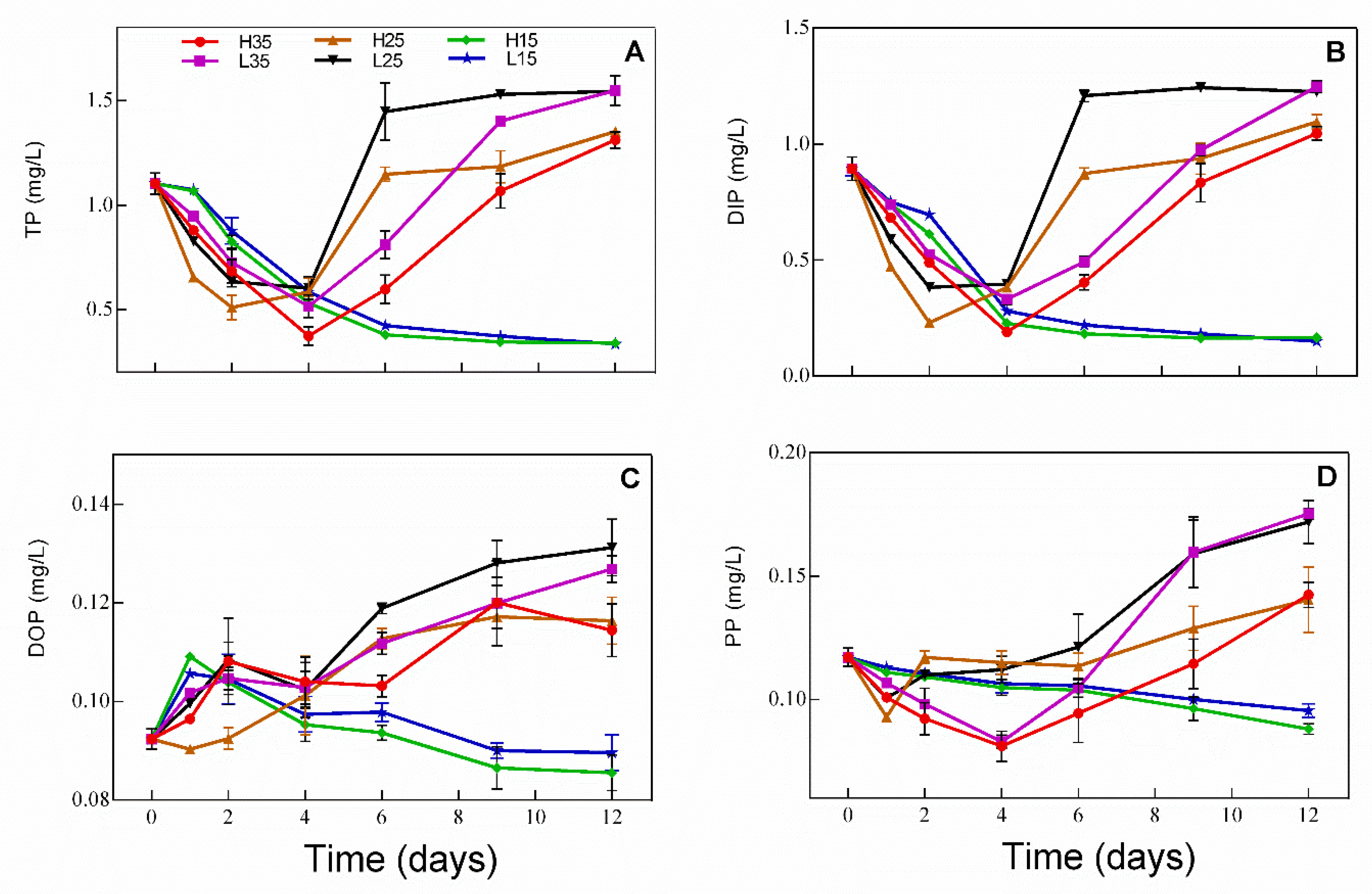
Concentration changes in different forms of phosphorus in water during the experiment are presented in Figure 2. TP (Figure 2A) and DIP (Figure 2B) show similar trends, with both decreasing over time in H15 and L15. TP and DIP decreased first and turned to an increasing trend on day 2 and day 4 in H25 and L25 and in H35 and L35, respectively. In those treatments, TP and DIP were even higher on day 12 than on day 0. DOP increased slightly in H15 and L15 on day 1 and then decreased gradually thereafter, while DOP increased gradually over time in the rest of treatments. Changes in PP were similar to that of TP, except that PP turned to increase on day 1 in H25 and L25.
3.3. Development of Periphyton
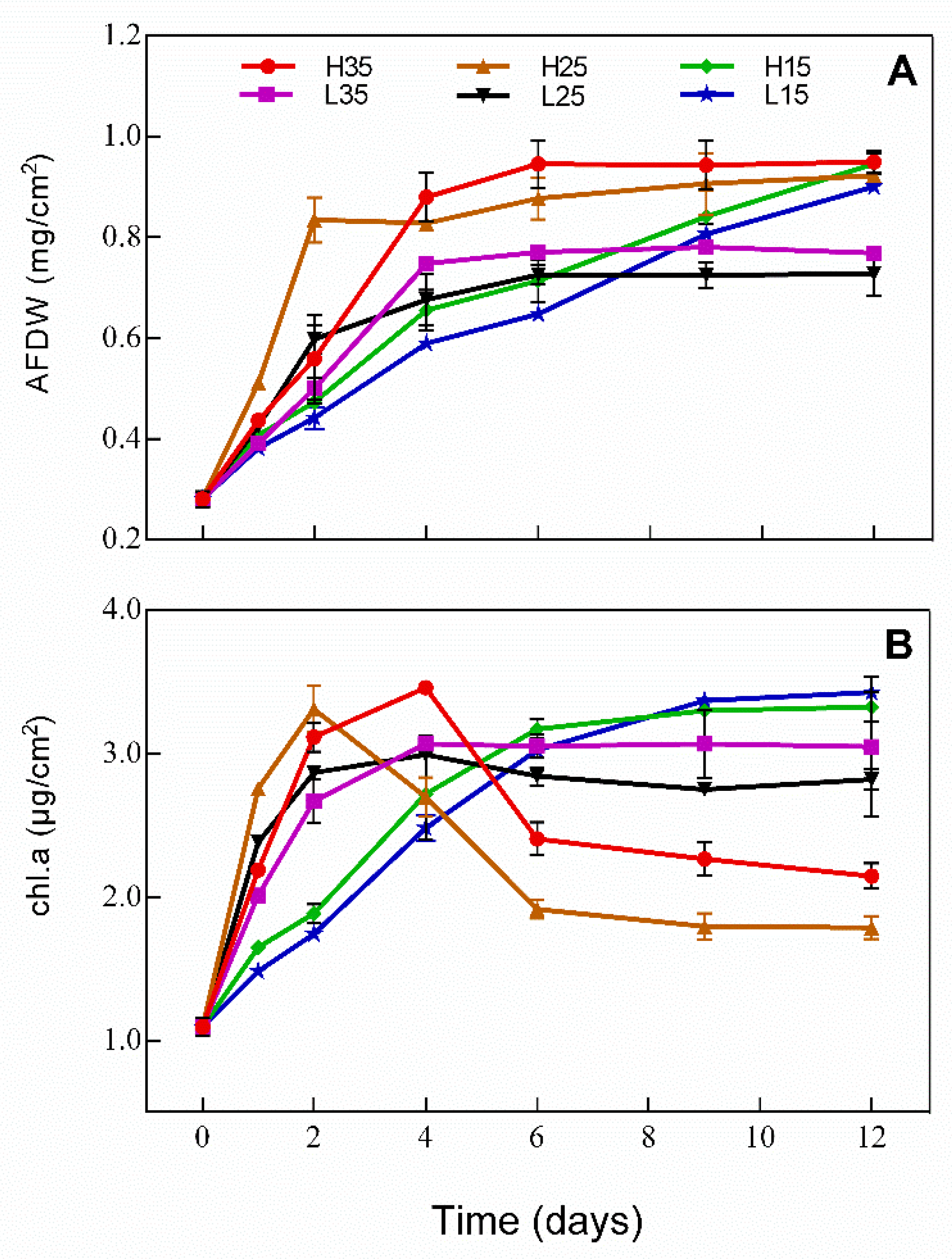
Changes in AFDW and Chl.a content of the periphyton is presented in Figure 3. AFDW increased over time in H15 and L15, while AFDW increased first and remained unchanged thereafter. On day 12, AFDW was the lowest in L25 and L35 and showed no difference in the rest of treatments. Chl.a showed similar trends to AFDW in H15, L15, L25, and L35. However, Chl.a increased first and then began to decrease in H25 and H35 on day 2 and day 4, respectively. On day 12, Chl.a was the highest in H15 and L15 and was the lowest in H25.
Algae composition of the periphyton was investigated on day 12 and algae belonging to 15 genera and 4 phyla were identified. As presented in Figure 4, algae composition was more diverse on day 0 with Lyngbya and Scenedesmus being the dominant genera. Algae composition became less diverse on day 12 in all treatments and Lyngbya became the only dominant genus in H35, L35, H25, and L25 while Lyngbya and Scenedesmus were dominant in H15 and L15 and were at a similar level.
3.4. Changes of Phosphorus and APase Activity in Periphyton
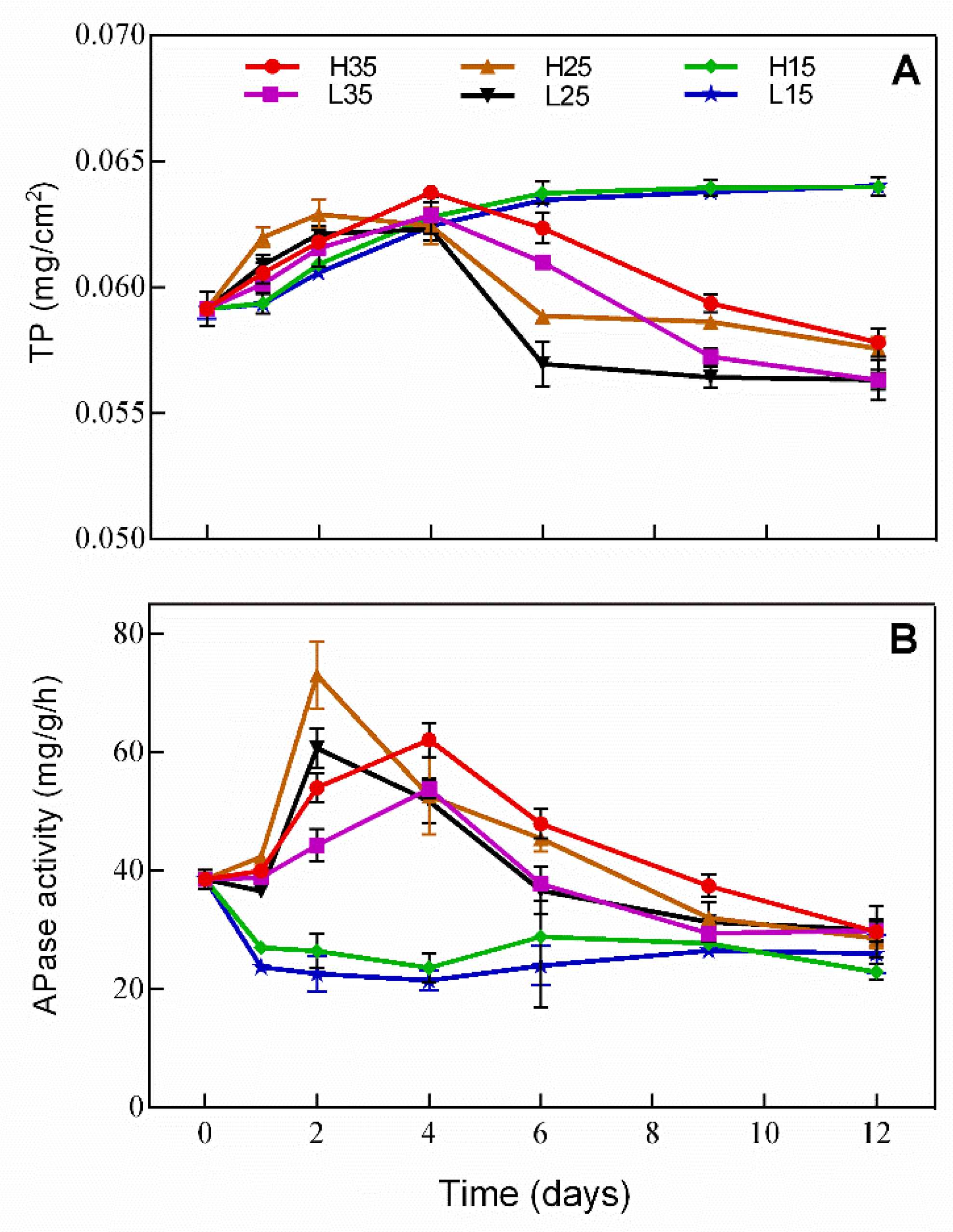
Changes in TP and APase activity in periphyton are presented in Figure 5. Changes in TP in periphyton show an opposite trend to that in water. TP increased over time in H15 and L15, but first increased and then decreased in the rest of the treatments, in which TP was even lower on day 12 than on day 1. APase activity in H15 and L15 decreased on day 1 and remained at a relatively low level thereafter, while in the rest of treatments, the trend of the APase activity was opposite to that of TP in water.
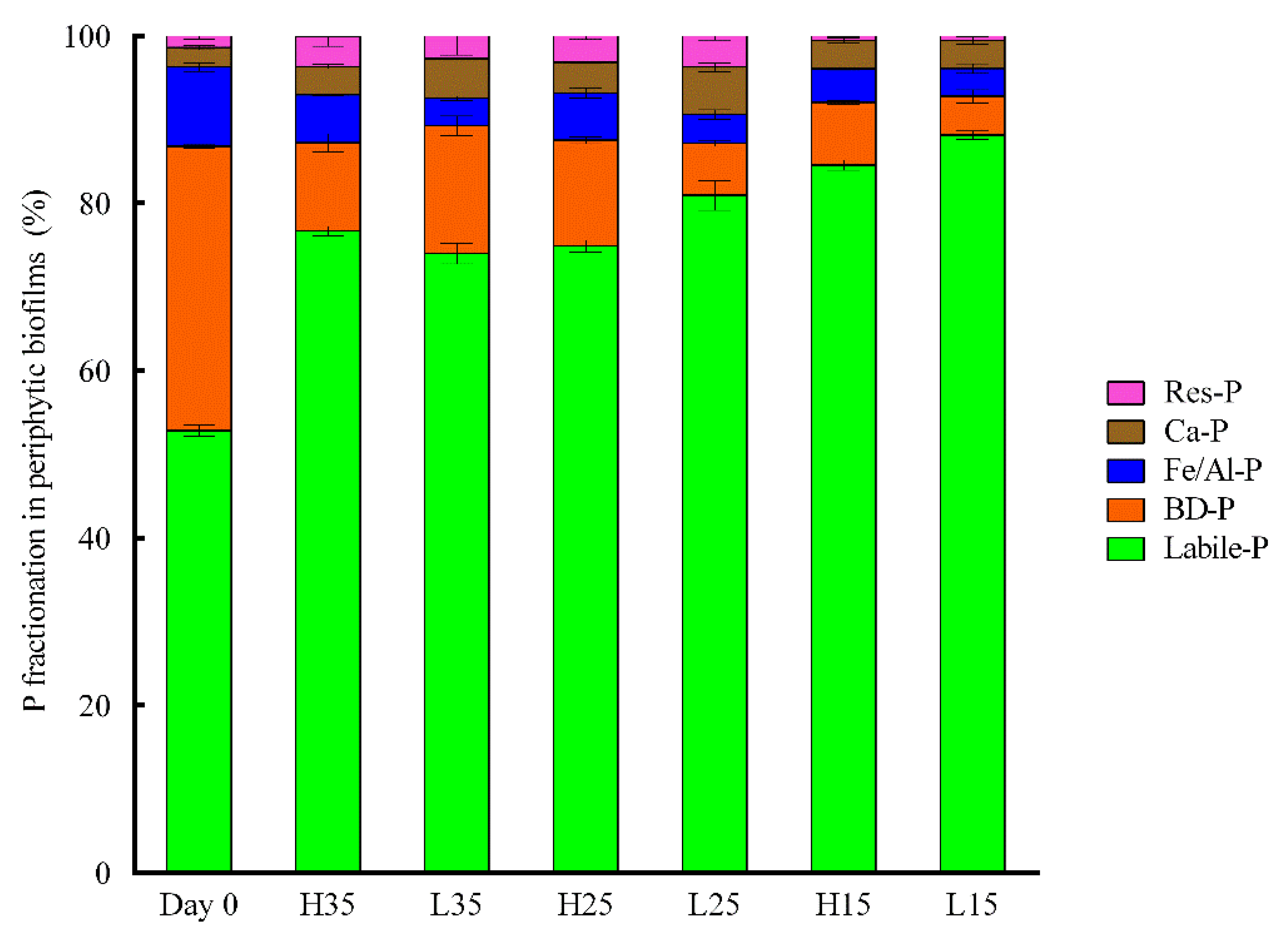
Fractionation of phosphorus is presented in Figure 6. Labile-P is the dominant fraction in all treatments. Compared to day 0, the proportion of labile-P increased in all treatments on day 12, while the proportion of BD-P and Fe/Al-P decreased. On day 12, periphyton in H15 and L15 have the highest labile-P fraction and the lowest Res-P.
3.5. Effects of Time and Treatments
The results of the repeated measurement ANOVA are summarized in Table 1. All parameters were significantly affected by time, illumination, and temperature. The interactive effects between time × illumination and time × temperature on all parameters were also significant for all parameters. However, the interactive effects of temperature x illumination on DO and APase activity and the interactive effects of time × illumination × temperature on pH and APA were not significant.
4. Discussion
In general, changes in pH and DO are used as important indicators of water chemistry and growth of aquatic organisms in water system. The DO was oversaturated and the pH was slightly basic in all treatments. This is related to the photosynthesis of algae in periphyton, which releases O2 and consumes CO2 during photosynthesis [13]. At 15 °C, DO and pH were relatively stable from day 1, suggesting that the photosynthetic activities can be maintained at a lower temperature, and the production and depletion of O2 reach an equilibrium. The decrease in DO and pH in the rest of the treatments after a few days suggests that the photosynthetic activities decreased and the activity of heterotrophic microorganisms was enhanced. As indicated in Figure 3B, Chl.a of the periphyton also decreased after few days, which is in accordance with the trend of DO. This is likely attributed to the senescence or the changes in the algae community structure in periphyton. Organic matters released during the decomposition of algae can stimulate the growth and activity of heterotrophic bacteria [20].
In closed water, phosphorus cannot be lost from the system. Comparing the initial concentration (D0) of phosphorus in the overlying water and the periphyton can be used as an indicator of phosphorus migration and transformation between them. Changes in phosphorus in the water and periphyton showed opposite trends. Phosphorus accumulated in periphyton over time at 15 °C, while phosphorus was firstly captured by periphyton and then released into the water after few days in other treatments, indicating that phosphorus can be retained in periphyton for a longer time under lower temperature conditions (15 °C). However, at 25 and 35 °C, periphyton turned from a sink to a source of phosphorus in closed water systems in a few days. Moreover, the initial rate of P captured by periphyton was greater and showed the greatest subsequent P release rate from day 4 to 6 in the 25 °C treatment (Figure 2). Higher temperatures can enhance the nitrification and denitrification processes in periphyton [21], which leads to a rapid depletion of nutrients for algae growth and initiates the senescence of periphyton. The depletion of phosphorus in water stimulated bacteria and algae produced APase. The APase activity increased in treatments at 25 and 35 °C and was maintained at low levels in treatments at 15 °C. The production of APase is an important process for microorganisms to deal with phosphorus limitation, which is responsible for the mineralization of organic phosphorus [22]. Many studies have showed that APase activity is affected by many factors such as pH, temperature, and substrate conditions [23]. The optimum pH was found to be around 9.5, which is close to the pH detected in this experiment, while lower temperatures generally inhibit APase activity [24,25].
Phosphorus capture in periphyton was more enhanced when illuminated at 70 than 20 μmol photons m−2 s−1 and similar trends were observed for treatments under the same temperature. However, a significantly higher amount of TP in periphyton was detected in treatments with higher illumination at 25 and 35 °C, which can be related to a higher biomass developed under higher illumination conditions, as reflected by AFDW (Figure 3A). However, TP in water and periphyton was not significantly different at 15 °C, suggesting that the effect of illumination on the development of periphyton was temperature dependent. This could be related to a different physiological acclimation of different algal species to different environmental conditions [26,27].
As illustrated in Figure 6, labile-P was the most abundant phosphorus fraction, suggesting that phosphorus captured in the periphyton can be easily released. The release of the phosphorus can be related to the disintegration of periphyton and the mineralization of organic phosphorus captured in periphyton [28]. An increase in DOP and PP for treatments at 25 and 35 °C toward the end of the experiment also reflect the disaggregation of periphyton. As showed in Table 1, time, illumination, and temperature had significant interactive effects on different forms of phosphorus. Therefore, periphyton in closed water systems act as a temporary storage for the introduced phosphorus and the transfer between water and periphyton changes overtime and was influenced by both illuminance and temperature conditions.
McCormick et al. [29] studied the role of periphyton in the Everglades Nutrient Removal Project and found that periphyton was an important short-term sink for phosphorus in treatment wetlands. They demonstrated that the retention of phosphorus is strongly related to the taxonomic composition of the periphyton, with cyanobacteria releasing more phosphorus back into the water. In our experiment, treatments at 25 and 35°C had a higher abundance of cyanobacteria at the end of the experiment (Figure 4), indicating that higher temperatures may favor the development of cyanobacteria-dominated periphyton and cause a quick shift from phosphorus capture to release. Drake et al. [30] studied the phosphorus immobilization and release in streams and demonstrated that phosphorus release occurred only in periphyton when light availability was low and the C:P ratio was also low. In open water systems such as streams, nutrients are continuously supplied and the microbial community can be maintained. In closed water systems, impulsive inputs of nutrients can temporarily stimulate the development of periphyton, but the rapid depletion of nutrients will cause the senescence of periphyton and release the captured phosphorus. Wu et al. [31] investigated the role of periphyton on phosphorus bioavailability in paddy fields and found that periphyton reduced phosphorus in water but increased phosphorus bioavailability for crops. The role of temporary phosphorus storage might contribute the increase of phosphorus bioavailability in paddy fields.
Periphyton can also have indirect effects on the phosphorus cycle. Although it has not been investigated here, previous works showed that periphyton can alter the environmental conditions at the water–sediment interface and subsequently affect the phosphorus retention and release in sediment [32,33]. Photosynthesis of periphyton can increase phosphate precipitation with calcium by increasing pH and promote phosphate adsorption by creating oxic conditions near the sediment surface [3]. However, decomposition and respiration of periphyton can lead to anoxia, which can release redox-sensitive phosphorus from the sediment [34]. Both direct and indirect effects lead to an initial decrease of phosphorus in water, followed by a release of phosphorus into the water from periphyton and sediment. This process is influenced by both temperature and illumination conditions, as shown in this work.
5. Conclusions
Results of this study reveal that periphyton act as a short-term sink of phosphorus in closed water systems following nutrient input. Phosphorus captured in periphyton can be released back to water following the depletion of nutrients, which causes the senescence of the periphyton. This process is strongly affected by illumination and temperature conditions and their interactions. Phosphorus can be retained in periphyton for a longer time at lower temperatures. Illumination enhanced phosphorus capture at higher temperatures. This work contributes to a better understanding of the role of periphyton in phosphorus cycling in closed water systems.
Author Contributions
Conceptualization, Y.Z. and C.W.; Methodology, Y.Z.; Validation, C.W., X.C. and X.X.; Formal analysis, Y.Z.; Investigation, Y.Z.; Resources, C.W.; X.C.; Data curation, Y.Z.; Writing—original draft preparation, Y.Z.; Writing—review and editing, C.W.; Supervision, X.C. and X.X.; Project administration, C.W.; Funding acquisition, C.W.
Acknowledgments
This work is supported by the State Key Development Program for Basic Research of China (2015CB158200) and the State Key Laboratory of Freshwater Ecology and Biotechnology (2019FBZ03).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Verdegem, M.; Van Dam, A.; Azim, M.; Beveridge, M. Periphyton ecology, exploitation and management: Knowledge gaps and directions for future research. In Periphyton: Ecology, Exploitation and Management; CABI Publishing: Wallingford, UK, 2005; pp. 299–307. [Google Scholar]
- Vadeboncoeur, Y.; Steinman, A.D. Periphyton function in lake ecosystems. Sci. World J. 2002, 2, 1449. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K. The role of periphyton in phosphorus retention in shallow freshwater aquatic systems. J. Appl. Phycol. 2003, 39, 840–849. [Google Scholar] [CrossRef]
- Ilker, U.; Ashvini, C.; Ogram, A.V. Cellulolytic, fermentative, and methanogenic guilds in benthic periphyton mats from the Florida Everglades. FEMS Microbiol. Ecol. 2010, 61, 337–347. [Google Scholar]
- Wolfe, J.E.; Lind, O. Phosphorus uptake and turnover by periphyton in the presence of suspended clays. Limnology 2010, 11, 31–37. [Google Scholar] [CrossRef]
- Boelee, N.; Temmink, H.; Janssen, M.; Buisman, C.; Wijffels, R. Nitrogen and phosphorus removal from municipal wastewater effluent using microalgal biofilms. Water Res. 2011, 45, 5925–5933. [Google Scholar] [CrossRef]
- Guzzon, A.; Bohn, A.; Diociaiuti, M.; Albertano, P. Cultured phototrophic biofilms for phosphorus removal in wastewater treatment. Water Res. 2008, 42, 4357–4367. [Google Scholar] [CrossRef]
- Sukačová, K.; Trtílek, M.; Rataj, T. Phosphorus removal using a microalgal biofilm in a new biofilm photobioreactor for tertiary wastewater treatment. Water Res. 2015, 71, 55–63. [Google Scholar] [CrossRef]
- Lu, H.; Feng, Y.; Wang, J.; Wu, Y.; Shao, H.; Yang, L. Responses of periphyton morphology, structure, and function to extreme nutrient loading. Environ. Pollut. 2016, 214, 878–884. [Google Scholar] [CrossRef]
- Trochine, C.; Guerrieri, M.E.; Liboriussen, L.; Lauridsen, T.L.; Jeppesen, E. Effects of nutrient loading, temperature regime and grazing pressure on nutrient limitation of periphyton in experimental ponds. Freshwater Biol. 2014, 59, 905–917. [Google Scholar] [CrossRef]
- Wagner, K.; Besemer, K.; Burns, N.R.; Battin, T.J.; Bengtsson, M.M. Light availability affects stream biofilm bacterial community composition and function, but not diversity. Environ. Microbiol. 2016, 17, 5036–5047. [Google Scholar] [CrossRef]
- Sanches, L.F.; Guariento, R.D.; Caliman, A.; Bozelli, R.L.; Esteves, F.A. Effects of nutrients and light on periphytic biomass and nutrient stoichiometry in a tropical black-water aquatic ecosystem. Hydrobiologia 2011, 669, 35–44. [Google Scholar] [CrossRef]
- Villanueva, V.D.; Font, J.; Schwartz, T.; Romaní, A.M. Biofilm formation at warming temperature: Acceleration of microbial colonization and microbial interactive effects. Biofouling 2011, 27, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Winkworth, C.L.; Salis, R.K.; Matthaei, C.D. Interactive multiple-stressor effects of the antibiotic monensin, cattle effluent and light on stream periphyton. Freshwater Biol. 2015, 60, 2410–2423. [Google Scholar] [CrossRef]
- Di Pippo, F.; Ellwood, N.T.W.; Guzzon, A.; Siliato, L.; Micheletti, E.; de Philippis, R.; Albertano, P.B. Effect of light and temperature on biomass, photosynthesis and capsular polysaccharides in cultured phototrophic biofilms. J. Appl. Phycol. 2012, 24, 211–220. [Google Scholar] [CrossRef]
- MEPC (Ministry of Environmental Protection of China). Water and Wastewater Monitoring and Analysis Methods; China Environ. Sci. Press: Beijing, China, 2002.
- Scott, J.T.; Lang, D.A.; King, R.S.; Doyle, R.D. Nitrogen fixation and phosphatase activity in periphyton growing on nutrient diffusing substrata: Evidence for differential nutrient limitation in stream periphyton. J. N. Am. Benthol. Soc. 2009, 28, 57–68. [Google Scholar] [CrossRef]
- Pei, G.; Wang, Q.; Liu, G. The role of periphyton in phosphorus retention in shallow lakes with different trophic status, China. Aquat. Bot. 2015, 125, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Sci. Press: Beijing, China, 2006. [Google Scholar]
- Grossart, H.P.; Czub, G.; Simon, M. Algae–bacteria interactions and their effects on aggregation and organic matter flux in the sea. Environ. Microbiol. 2006, 8, 1074–1084. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiong, X.; Wu, C.; Xia, Y.; Li, J.; Wu, Y. Influence of light and temperature on the development and denitrification potential of periphytic biofilms. Sci. Total Environ. 2018, 613, 1430–1437. [Google Scholar] [CrossRef]
- Duport, J.P. Responses of lotic periphyton to pulses of phosphorus: P-flux controlled growth rate. Freshwater Biol. 2012, 57, 2602–2612. [Google Scholar]
- Verchot, L.V.; Borelli, T. Application of para -nitrophenol (pNP) enzyme assays in degraded tropical soils. Soil Biol. Biochem. 2005, 37, 625–633. [Google Scholar] [CrossRef]
- Hui, D.; Mayes, M.A.; Wang, G. Kinetic parameters of phosphatase: A quantitative synthesis. Soil Biol. Biochem. 2013, 65, 105–113. [Google Scholar] [CrossRef]
- Qian, Y.; Liang, X.; Chen, Y.; Lou, L.; Cui, X.; Jie, T.; Li, P.; Cao, R. Significance of biological effects on phosphorus transformation processes at the water–sediment interface under different environmental conditions. Ecol. Eng. 2011, 37, 816–825. [Google Scholar] [CrossRef]
- Kumar, D.; Kvíderová, J.; Kaštánek, P.; Lukavský, J. The green alga Dictyosphaerium chlorelloides biomass and polysaccharides production determined using cultivation in crossed gradients of temperature and light. Eng. Life Sci. 2017, 17, 1030–1038. [Google Scholar] [CrossRef]
- Wu, H.; Gao, G.; Zhong, Z.; Li, X.; Xu, J. Physiological acclimation of the green tidal alga Ulva prolifera to a fast-changing environment. Mar. Environ. Res. 2018, 137, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Halvorson, H.M.; Scott, E.E.; Entrekin, S.A.; Evans-White, M.A.; Scott, J.T. Light and dissolved phosphorus interactively affect microbial metabolism, stoichiometry and decomposition of leaf litter. Freshwater Biol. 2016, 61, 1006–1019. [Google Scholar] [CrossRef]
- Mccormick, P.V.; Iii, R.B.E.S.; Chimney, M.J. Periphyton as a potential phosphorus sink in the Everglades Nutrient Removal Project. Ecol. Eng. 2006, 27, 279–289. [Google Scholar] [CrossRef]
- Drake, W.M.; Scott, J.T.; Haggard, B.; Sharpley, A.; Rogers, C.W.; Grantz, E.M. The effect of periphyton stoichiometry and light on biological phosphorus immobilization and release in streams. Limnology 2012, 13, 97–106. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Lu, H.; Wu, C.; Kerr, P. Periphyton: An important regulator in optimizing soil phosphorus bioavailability in paddy fields. Environ. Sci. Pollut. Res. 2016, 23, 21377–21384. [Google Scholar] [CrossRef]
- Lu, H.; Wan, J.; Li, J.; Shao, H.; Wu, Y. Periphytic biofilm: A buffer for phosphorus precipitation and release between sediments and water. Chemosphere 2016, 144, 2058–2064. [Google Scholar] [CrossRef]
- Zhang, X.F.; Mei, X.Y. Effects of benthic algae on release of soluble reactive phosphorus from sediments: A radioisotope tracing study. Water Sci. Eng. 2015, 8, 127–131. [Google Scholar] [CrossRef]
- Wood, S.A.; Depree, C.; Brown, L.; Mcallister, T.; Hawes, I. Entrapped Sediments as a Source of Phosphorus in Epilithic Cyanobacterial Proliferations in Low Nutrient Rivers. PLoS ONE 2015, 10, e0141063. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Changes in (A) DO and (B) pH in water in different treatments during the experiment.

Figure 2.
Concentration changes in the different forms of phosphorus ((A) TP, (B) DIP, (C) DOP, and (D) PP) in water in different treatments during the experiment.
Figure 2.
Concentration changes in the different forms of phosphorus ((A) TP, (B) DIP, (C) DOP, and (D) PP) in water in different treatments during the experiment.

Figure 3.
Changes in (A) ash-free dry weight (AFDW) and (B) Chlorophyll a (Chl.a) of the periphyton in different treatments during the experiment.
Figure 3.
Changes in (A) ash-free dry weight (AFDW) and (B) Chlorophyll a (Chl.a) of the periphyton in different treatments during the experiment.

Figure 4.
Algae composition of the periphyton on day 0 and day 12 in different treatments.

Figure 5.
Changes in (A) TP and (B) APase activity in periphyton in different treatments during the experiment.
Figure 5.
Changes in (A) TP and (B) APase activity in periphyton in different treatments during the experiment.

Figure 6.
Phosphorus fractionation in the periphyton on day 0 and day 12 in different treatments.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results (p values) of repeated measurement ANOVA. Illumination (I), time (day), and temperature (T).
Table 1.
Results (p values) of repeated measurement ANOVA. Illumination (I), time (day), and temperature (T).
| Source | df | pH | DO | AFDW | Chl.a | TP | DIP | DOP | PP | APase Activity |
|---|---|---|---|---|---|---|---|---|---|---|
| Day | 6 | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * |
| I | 1 | 0.000 * | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.000 * | 0.024 * | 0.000 * | 0.000 * |
| T | 2 | 0.000 * | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * |
| Day × I | 6 | 0.000 * | 0.001 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * |
| Day × T | 12 | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * |
| T× I | 2 | 0.001 * | 0.231 | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.001 * | 0.000 * | 0.053 |
| Day × I × T | 12 | 0.052 | 0.024 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.034 * | 0.001 * | 0.45 |
Note: * indicates significant difference at 95% confidence interval.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, Y.; Chen, X.; Xiong, X.; Wu, C. Capture and Release of Phosphorus by Periphyton in Closed Water Systems Influenced by Illumination and Temperature. Water 2019, 11, 1021. https://doi.org/10.3390/w11051021
AMA Style
Zhao Y, Chen X, Xiong X, Wu C. Capture and Release of Phosphorus by Periphyton in Closed Water Systems Influenced by Illumination and Temperature. Water. 2019; 11(5):1021. https://doi.org/10.3390/w11051021
Chicago/Turabian StyleZhao, Yanhui, Xiaofei Chen, Xiong Xiong, and Chenxi Wu. 2019. "Capture and Release of Phosphorus by Periphyton in Closed Water Systems Influenced by Illumination and Temperature" Water 11, no. 5: 1021. https://doi.org/10.3390/w11051021
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.