Prokaryotic Community Composition Affected by Seasonal Changes in Physicochemical Properties of Water in Peat Bog Lakes


1
Department of Microbiology and Mycology, University of Warmia and Mazury in Olsztyn, Oczapowskiego str. 1a, 10-719 Olsztyn, Poland
2
Department of Water Resources, Climatology and Environmental Management, University of Warmia and Mazury in Olsztyn, Plac Łódzki 2, 10-719 Olsztyn, Poland
3
Environmental Biotechnology Department, Silesian University of Technology, Akademicka 2, 44-100 Gliwice, Poland
*
Author to whom correspondence should be addressed.
Water 2018, 10(4), 485; https://doi.org/10.3390/w10040485
Submission received: 16 February 2018
/
Revised: 11 April 2018
/
Accepted: 12 April 2018
/
Published: 16 April 2018
(This article belongs to the Special Issue Freshwater Quality as a Driver of Aquatic Ecosystem Health)
Abstract
:Based on a three-year study on the prokaryotic community composition in peat bog lakes surrounded by a floating mat of Sphagnum sp. moss in the conditions of Northeast Poland (Central Europe), we verified the relationship between 20 water parameters and main Eubacteria and Archaea phyla for specific sites: the subsurface (pelagic zone), near-bottom (benthic zone), and the Sphagnum mat (ecotone zone). Abundance and composition of the main aquatic bacteria phyla (Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Deltaproteobacteria, Actinobacteria, and Cytophaga-Flavobacteria) and Archaea were associated with different combinations of physico-chemical parameters of water, and followed temporal variations of temperature, dissolved organic carbon (DOC), aromaticity, and water color. Redundancy analysis (RDA) showed that water acidity is a less significant predictor of bacterial activity; however, we have found Betaproteobacteria negatively correlated (r = −0.49, p = 0.01), while Actinobacteria positively correlated (r = 0.21, p = 0.05) to pH. This relation was the most significant in the ecotone zone. In the overall bacteria community structure Betaproteobacteria dominated (18.3%) regardless of site or season, except for winter when, at low temperatures and DOC concentrations, Actinobacteria increased to 22.9%. The Archaea fraction was uniform (11%) in seasons and sites and showed no specific preferences to physico-chemical predictors. Although the water parameters from the Sphagnum mat did not differ significantly from pelagic water, its role as a source of allochthonous organic matter is crucial for bacteria activity. The relations between peat bog lake attributes and seasonal changes in bacterial diversity demonstrated a distinct divergent pattern for each prokaryote. Obtaining results will provide support for any future evaluation of the effects of environmental variables on prokaryotic community structures in peat bog lakes.
Keywords:
prokaryotic community composition; humic lakes; peat bog; pelagial; benthic zone; ecotone; water quality; RDA1. Introduction
Peat bog lakes, usually surrounded by Sphagnum-dominated peatland, create one of the most extensive types of wetlands in the temperate climate zone, characterized by high acidity, water color, and lower concentrations of mineral compounds, but higher concentration of organic matter when compared to eutrophic or oligotrophic lakes [1]. These ombrotrophic ecosystems are decoupled from groundwater of the surrounding watershed and receive all water and nutrient inputs from the atmosphere.
Understanding the composition and the dynamics of the microbial community is crucial because microorganisms are known to play key roles in many biogeochemical processes. In peat bog lakes, similarly to other types of freshwater ecosystems, microorganisms play a fundamental role in the decomposition of organic matter and system respiration and, thus, they recycle nutrients in a complex manner involved in various geochemical cycles as the nitrogen, phosphorus, and carbon cycles [2]. Whereas the seasonal dynamics of microorganisms in eutrophic and oligotrophic ecosystems have been well recognized [3], the diversity and population dynamics of bacterioplankton in humic lakes are still poorly understood. The unique feature of the humic lakes is the limited availability of nutrients, what makes other factors significant in the regulation of bacterial community composition (BCC), for example, the binding of metals and organic compounds affects their bioavailability and natural acidity [4].
For this reason, peat bog lakes are considered as specific environments, and due to their high natural value, understanding the structure of microbial communities is crucial for the ecosystem in which bacteria make it impossible to maintain homeostasis.
Individual bacterial populations are highly dynamic and can differ strongly both in season and water depth within an individual ecosystem in terms of the response to resource availability (in particular, organic carbon and inorganic nutrients, such as nitrogen and phosphorus), but also in terms of local environmental conditions, as acidity, temperature, or light availability [5] that, in turn, determines the food web structure. Long-term monitoring showed that aquatic systems of this type undergo fairly predictable variations in physical parameters and resource availability on an annual scale [6]. Yannarell [7] showed that lakes experience quite dramatic changes in BCC within- and between-years, regardless of lake trophic status.
In freshwater ecosystems, both the quality and quantity of organic matter, which is predominantly composed of dissolved organic matter, have been shown to control the metabolic activity and composition of microbial organisms [8]. This relationship was confirmed by Findlay [9], who observed increased metabolic activities accompanied by changes in the benthic microbial community at presence of highly-bioavailable organic matter. So far, heterotrophic bacteria have been considered as the main component of planktonic and benthic carbon cycling in freshwater ecosystems as they use organic compounds as a source of carbon [10].
Microbial processing and photodegradation (solar UV) of dissolved organic matter is an important source of organic matter in sediments, together with sinking particulate organic matter (POM) [11]. Dissolved organic matter can originate from internal primary production within an ecosystem, i.e., autochthonous organic matter, or from terrestrial inflow and precipitation, i.e., allochthonous organic matter. In most lacustrine ecosystems, allochthonous DOM predominates over autochthonous DOM [12]. In case of peat bog lakes, the ecotone zone, built of Sphagnum moss cover, plays the role of a direct supplier of allochthonous DOM. The amount of carbon sequestered in net primary production, mainly by Sphagnum mosses, exceeds the amount of carbon lost by decomposition of organic matter by microorganisms [13]. Ecotone contributes to the increased dissolved organic carbon (DOC) concentrations and acidification of water [14]. In this way, the ecotone alters microbial OM processing [5] and the structure of bacterial communities [15].
Several studies have reported on the abundance and productivity of bacterial communities in humic lakes [3,16,17,18], but few studies have examined factors that contribute to BCC changes in various ecological parts of a single lake system [19] A focus on identifying dynamic and persistent bacterial populations should lead to a greater understanding of factors influencing bacterial community structure in humic lakes [3,19]. To do so, we designed a multiyear study of microbial populations in three small lakes located in the protected area within the Wigry National Park, located in Northeastern Poland. The study required a spatio-temporal approach to the investigation of prokaryotic community composition in relation to hydrochemical predictors in three integral intra-system habitats: (i) the subsurface pelagic zone; (ii) the near-bottom zone; and (iii) the waterlogged Sphagnum mat, hereafter called the ecotone zone, which is a transition zone between the peat bog lake and the mineral surroundings. The microbial ecology of humic lakes requires, however, not only the study of microorganisms, per se, but, because of the multidisciplinary approach of the various interactions of the microorganisms, it should produce knowledge on abiotic factors that shape the microbial community, regulating both its quantitative composition and diversity, and affecting the seasonal dynamics of bacterioplankton [20].
For this purpose, we followed quantitative changes in the bacterial community composition and in the physicochemical characteristics of the pelagic, benthic, and ecotone zones of three natural dystrophic lakes located in the Wigry National Park (Northeast Poland). In the present study, we are going to determine the composition of bacteria in peat bog lakes, as well as identify linkages between the bacterial community composition in different parts of the lake ecosystems and key environmental (hydrochemical) parameters of water. The obtained results should verify two research hypotheses based on the quality and quantity of prokaryotic communities: (i) the Sphagnum mat ecotone creates an intrinsic habitat in peat bog lakes, which is a main source of organic matter; and (ii) peat bog lakes, characterized by advanced processes of dystrophy, create habitats suitable for the specific structure of bacterioplankton with a low diversity of prokaryotes, not favorable to bacterioplankton productivity.
2. Materials and Methods
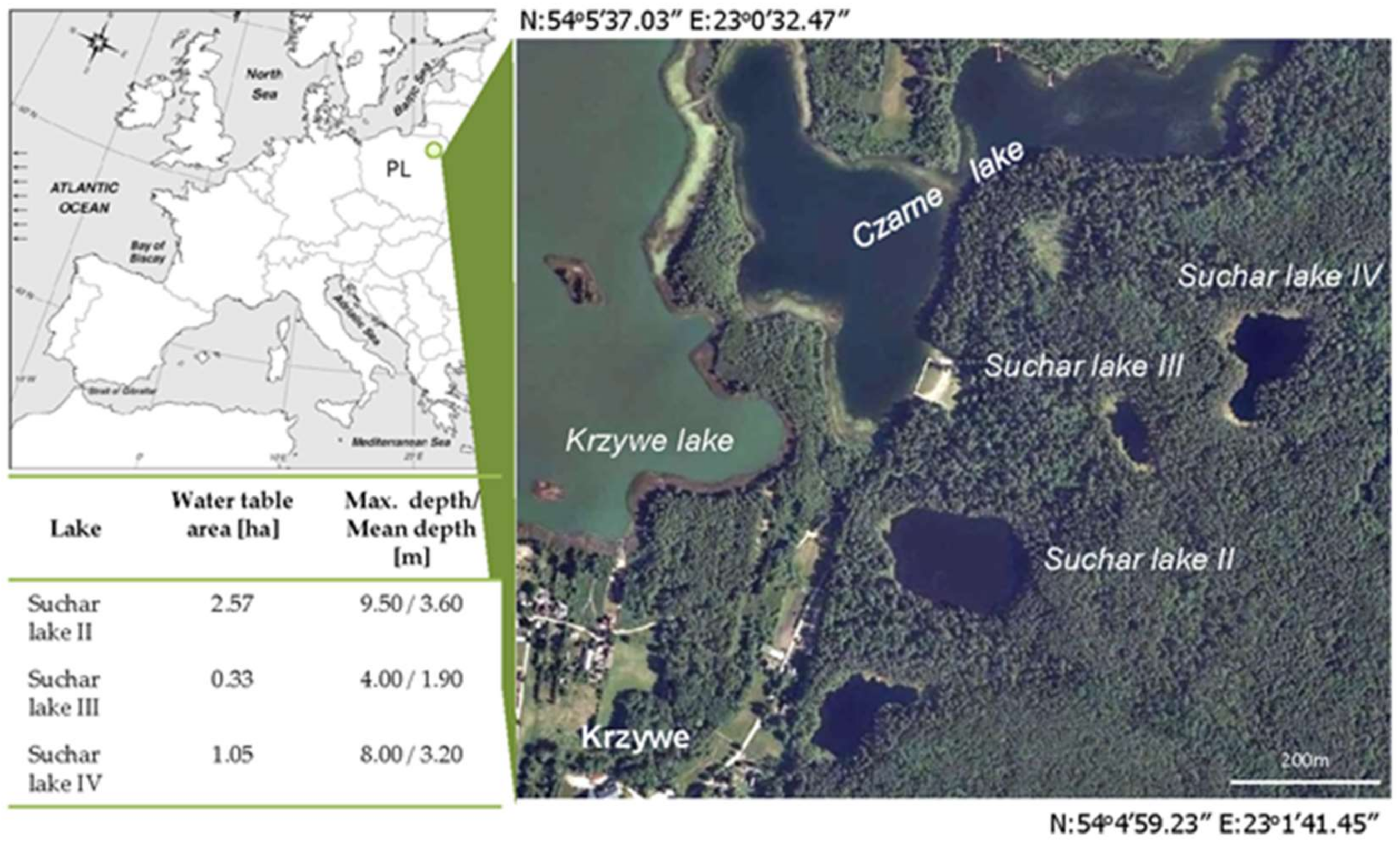
From among 18 ‘Suchar’ lakes located in the Wigry National Park in Northeastern Poland, we selected for the present study three small dystrophic lakes called Suchar lake II, Suchar lake III, and Suchar lake IV (Figure 1). The chosen lakes are located in the northwest of Wigry Lake, the largest lake in the area. They are small, surrounded by floating mats of Sphagnum mosses, which measure from 0.50 to 3.00 ha in area. They are in the intermediate stage of their ontogeny.
Their maximal depths do not exceed 8.00 m, while an average depths range from 1.90 to 3.60 m. The development and relationship to the hydrology of these sites is attributed to the remnants of the glacial period.
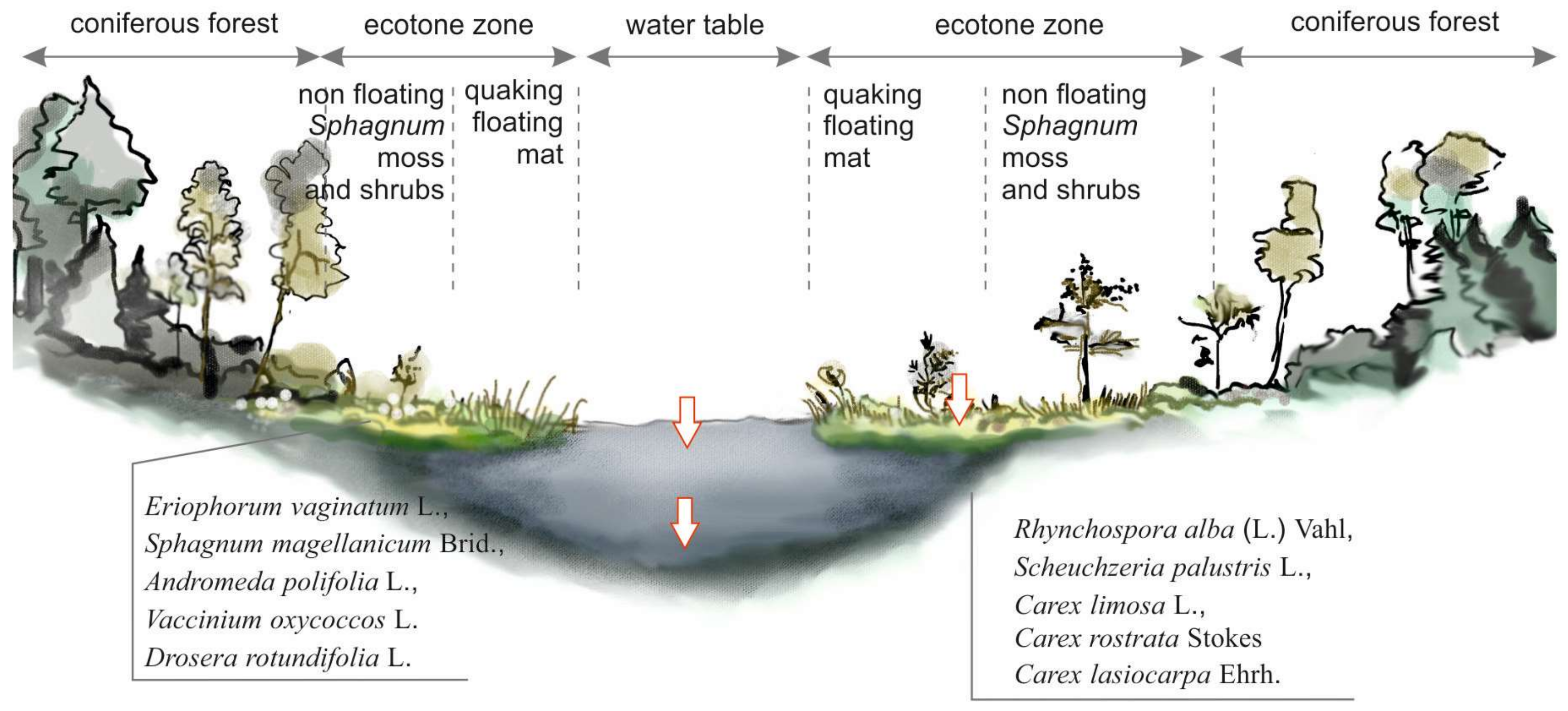
The water bodies are outflowless, dark-brown in water color, acidified, characterized by low concentrations of calcium, and have a high content of humic acids supplying the lakes from their catchments overgrown by the bog pine forest (Vaccinio uliginosi-Pinetum) or oak-hornbeam forests (Tilio-Carpinetum). Forests adjacent to the lakes protect them against winds and overshadows their water tables, thus, the ‘Suchar lakes’ belong to stratified and poorly-mixed water bodies. The wet area on the outer margins, known as a moat or lagg, is adjacent to the upland slopes. Vegetation at these sites is dominated by Sphagnum mosses and low shrubs typical of northern bogs, including Andromeda polifonia, Ledum palustre, Oxycoccus palustris, Menyanthes trifoliata, Eriophorum vaginatum, and Drosera sp. (Figure 2).
2.1. Physico-Chemical Parameters of Water
Water samples for physico-chemical analyses were taken simultaneously with bacteria sampling, during three years in four seasons: in spring, when water surface temperature was over 10 °C, in summer (t > 20 °C), fall (t < 15 °C), and in winter from under the ice cover. Water samples were taken in the central part (pelagic) of the lake from the subsurface and near-bottom layers, as well as from the ecotone zone, a moat—the hydrated part on the border area between the Sphagnum moss and boggy pine forest (Figure 2).
In situ measurements of dissolved oxygen (DO), temperature, pH, electrolytic conductivity (EC), and chlorophyll-a (chl-a) were performed using the YSI 6600R2™ calibrated multiprobe (Yellow Springs, OH, USA). Water transparency was measured with Secchi disc (20 cm in diameter). Water color was measured with the spectrophotometer DR/2010 (HACH, Loveland, CO, USA) using the APHA platinum-cobalt standard method, the attenuated radiation method (direct reading), and the HACH sulfide test, respectively [21]. The levels of TOC (total organic carbon), DOC (dissolved organic carbon), POC (particulate organic carbon), TN (total nitrogen), TDN (total dissolved nitrogen), N-NO3, N-NH4, TP (total phosphorus), and P-PO4 were measured with a spectrophotometer (Shimadzu UV–VIS 1601, Shimadzu Europa GmbH, Duisburg, Germany) and a Shimadzu TOCV-CSH total organic carbon analyzer [22]. Additionally, the DOC-specific UV absorbance at 254 nm (SUVA) was used as a proxy for the degree of aromaticity of DOM. All samples were analyzed in triplicate.
2.2. Microbial Communities Analysis
The total number of bacteria (TBN) was determined using epifluorescence microscopy [23]. Triplicate subsamples were fixed with neutralized formaldehyde (pH 7.4) at a final concentration of 4%. In the laboratory the samples were stained with DAPI (final concentration 1 μg mL−1) for 15 min in the dark, and gently filtered through 0.2 μm black Nuclepore filters. The bacteria were counted using an Olympus BX41 epifluorescence microscope. More than 1000 bacterial cells were counted in 20 fields of vision.
Procaryotic Community Composition: The community composition of bacterioplankton was analyzed for the 16S rRNA gene using denaturing gradient gel electrophoresis (DGGE), and fluorescent in situ hybridization DOPE-FISH (double labelling of oligonucleotide probes).
PCR-DGGE: Community genomic DNA was extracted within 12 h of sample collection. DNA was extracted according to the method described by [24]. To remove PCR inhibitors from the DNA samples an anti-inhibitor kit (A&A Biotechnology) was used. PCR amplification was performed on the 16S rRNA gene fragment using universal bacterial primers: 338f-GC 5′ CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GCC TAC GGG AGG CAG CAG 3′ and 518r-5′ ATT ACC GCG GCT GCT GG 3′ [25]. PCR was performed in a C1000 thermal cycler (Biorad), as previously described [26]. PCR products were separated by denaturing gradient gel electrophoresis (DGGE) in 8% (v/v) polyacrylamide gel with a 30–60% denaturant gradient (urea, Fluka). The separation was performed using the Dcode Mutation Detection System (BioRad) with a 1 TAE buffer (Tris, acetic acid, EDTA, pH 8.0) for 16 h at 60 °C and 40 V. DGGE gel was stained with SYBR Gold (1:10,000, Invitrogen) for 30 min, washed in MilliQ water for 30 min, and visualized and photographed under UV light with Quantity One 1D Software (Biorad). The same software was used for densitometric analysis of the obtained fingerprints. Due to the difficulties with sufficient PCR product obtainment for all the samples (the water samples contained large amount of PCR inhibitors and they were difficult to work with), the weakest fingerprints were used to estimate the densitometric threshold. On the basis of densitometric data, Shannon’s biodiversity index (H’) was calculated [27].
DOPE-FISH: Samples for the analysis of the microbial community structure were fixed with freshly-prepared buffered paraformaldehyde (pH 7.4) to a final concentration of 2% (v/v) and stored for several hours at 4 °C. The samples were filtered through 0.2 μm white polycarbonate filters, rinsed with sterile water, dried at room temperature, and stored at −20 °C. The structure of the bacterial community was investigated with fluorescence in situ hybridization (FISH) [28,29], with a modification described by Stoecker [30]. The bacterial community was analyzed with the use of probes (details are presented in Supplementary Materials Table S1): the EUB338I-III oligonucleotide probe targeting total bacteria [31], and the ARCH915 probe targeting Archaea [32]. Within the bacteria, the main taxonomic groups were determined using ALF968, BET42a, GAM42a, and DELTA 495abc probes for Alpha-, Beta-, Gamma-, and Deltaproteobacteria [33,34]; a CF319a probe for Cytophaga-Flavobacteria [33]; and an HGC69a probe for Actinobacteria [35]. Autofluorescence and non-specifically stained cells were determined with the NON338 negative control probe [36]. All oligonucleotide probes were double labelled with Cy3 dye [30]. Bacterial cells on the filter sections were observed with an epifluorescence microscope equipped with filter sets for DAPI and CY3. The fractions of DOPE-FISH stained bacteria in at least 1000 DAPI-stained cells per sample were quantified in triplicate.
Peat bog lake samples were taken in triplicate to determine the variability of DAPI counts. Probe-specific cell counts are presented as the percentage of cells visualized by DAPI.
2.3. Data Analyses
Before the statistical analyses, the datasets were log(x + 1) transformed, in order to stabilize variance. Normality was checked with the use of the Kolmogorov–Smirnov test. One-way analysis of variance (ANOVA) with the Tukey’s test, as a post-hoc procedure, was used to test differences in bacterial abundance and water chemistry parameters measured at three sites: the Sphagnum mat surface (ecotone zone), the lake subsurface layer (pelagic zone), and the near-bottom layer (benthic zone). We also run two-way ANOVA to check the differences among seasons and sites. The correlation analysis, mean abundances, and standard deviations (±SD) were calculated using Dell™ Statistica™ 13.1.
The response of the microbiological communities to environmental variables was analyzed using multivariate statistical analyses. This method was chosen after initial analysis by detrended correspondence analysis (DCA) revealed that the data exhibited a unimodal, rather than a linear, response to the environmental variables. The DCA of the microbiological parameters was used to determine whether linear or unimodal ordination methods should be applied [37].
DCA was used first to determine the variability in the studied assemblages: if a gradient length was over 4 SD, species in the data show a clear unimodal response along the gradient. The gradient length amounted to SD = 0.054, which indicated a linear variation, providing justification for the further use of redundancy analysis (RDA), which is a direct gradient analysis that summarizes the relations between bacteria classes and water quality parameters. The dataset was centered and standardized by species, due to the different units of environmental variables. To rank the importance of the individual explanatory variable, automatic forward selection of environmental variables was used. Before each addition, the explanatory effect of the candidate variable was evaluated using the Monte Carlo permutation full-model test with 999 unrestricted permutations [38]. DCA and RDA were performed using CANOCO 4.5. Finally, to represent biplots, we used the program CANODRAW within the CANOCO package.
To determine the associations between the groups of microorganisms and environmental factors (season x site) we performed a two way cluster analysis (TWCA). Standardized data were analyzed with the Bray–Curtis distance measure and flexible beta (=−0.25) linkage method using PC_ORD 6.08 software [39].
3. Results
3.1. Physico-Chemical Parameters of Water
Water in the studied peat bog lakes was brown, nutrient-poor, humic, acid rich, and lime-free (dystrophic). The studied benthic, pelagial, and ecotone zones in the humic lakes demonstrated significant differences in values of water quality parameters (Table 1).
The pH of the water in the studied lakes was very stable over the course of the study and its median value amounted to 4.38. No statistically significant differences (Tukey test, one-way ANOVA, p < 0.05) in terms of pH between the studied lakes sites was stated. The lowest pH (3.63 in the pelagic zone in spring) and the darkest water color (1306.67 mg∙Pt∙L−1 at the bottom in spring) were found in Suchar Lake IV, which is surrounded by the well-developed and largest zone of moat. In other lakes darker water color was characteristic for the bottom layer. The highest pH of 5.70 was noted in the pelagic zone in Suchar II and Suchar IV during summer. The lakes were generally abundant in organic matter where DOC constituted 89% of TOC content (23.4 mg∙L−1). Electrolytic conductivity, as an indirect measure of water mineralization, showed relatively low values in all sites (<30 µS∙cm−1) when compared to, e.g., eutrophicated water bodies [40]. However, EC values for the bottom zone were significantly higher (37 µS∙cm−1) compared to pelagic or ecotone zones (Tukey test, one-way ANOVA, p < 0.05).
The Suchar lakes receive large inputs of organic matter from the extensive Sphagnum mat that surrounds the water table. The terrestrial inputs of DOC are supplemented by autochthonous production of organic matter, mainly during the ice-free period. As a result, the studied lakes showed elevated levels of DOC, the averages of which ranged from 15.6 to 29.2 mg∙C∙L−1. DOC values were significantly higher in fall, when mean values amounted to 36.80 mg∙C∙L−1, and then in winter declined to 8.79 mg∙C∙L−1. The accumulation of humic compounds and decomposed organic matter from adjacent peatlands significantly contributed to an increase of dissolved carbon in benthic zone.
The aromatic fraction, SUVA254, was nearly uniform across the studied lakes (no statistical differences among sites, Tukey test, ANOVA, p < 0.05) and varied from 3.7–4.2 L∙mg−1∙m−1, which is indicative for humic lakes characterized by high DOC concentrations of allochthonous origin, acidic pH, and high content of humic substances.
N cycling in the studied lakes was dominated by organic forms which constituted as much as three-quarters of the N budget. In the subsurface layer of the lakes, a high concentration of nitrate nitrogen compared to ammonia nitrogen was found. The average concentration of N-NO3 in the surface layer of the group of Suchar lakes amounted to 0.16 mg∙L−1, while N-NH4 amounted to 0.03 mg∙L−1. The reverse situation took place at the bottom, where concentrations of nitrate nitrogen were noticed at the level of 0.07 mg∙L−1, while the concentrations of N-NH4 amounted on average to 0.61 mg∙L−1. Ammonia concentrations increased by a factor of 20 from the surface to the bottom at all three sites, and did not substantially differ between the lakes. Sedimentation of the matter also favored accumulation of phosphorus, concentrations of which were the highest at the bottom. The contribution of organic fraction (TOP) in TP accumulated in the bottom amounted to 54%, while the share of TOP in the pelagic zone increased to 60%, and that in the ecotone zone achieved 85%. The highest levels of both TN and TP were also found in the bottom layer, particularly TN, where concentrations reached the maximum level in spring.
Overall productivity of the ecosystems was related to the concentrations of chl_a, which, in the benthic zone, was three times higher (~109 μg∙L−1) as in the ecotone or pelagic zones (ca. 28–32 μg∙L−1).
All the studied parts of the peat bog lakes experienced high concentrations of SIO2 during late winter–early spring months. The increase in SiO2 reflects diatom activity in various aquatic ecosystems in climatic conditions of Northeast Europe, which results in water saturation with DO. Ice cover favors significant diatom development, particularly in the subsurface layer of water, which promotes elevated silica content, both in lake water and bottom sediments.
3.2. Bacterial Abundance and Community Composition
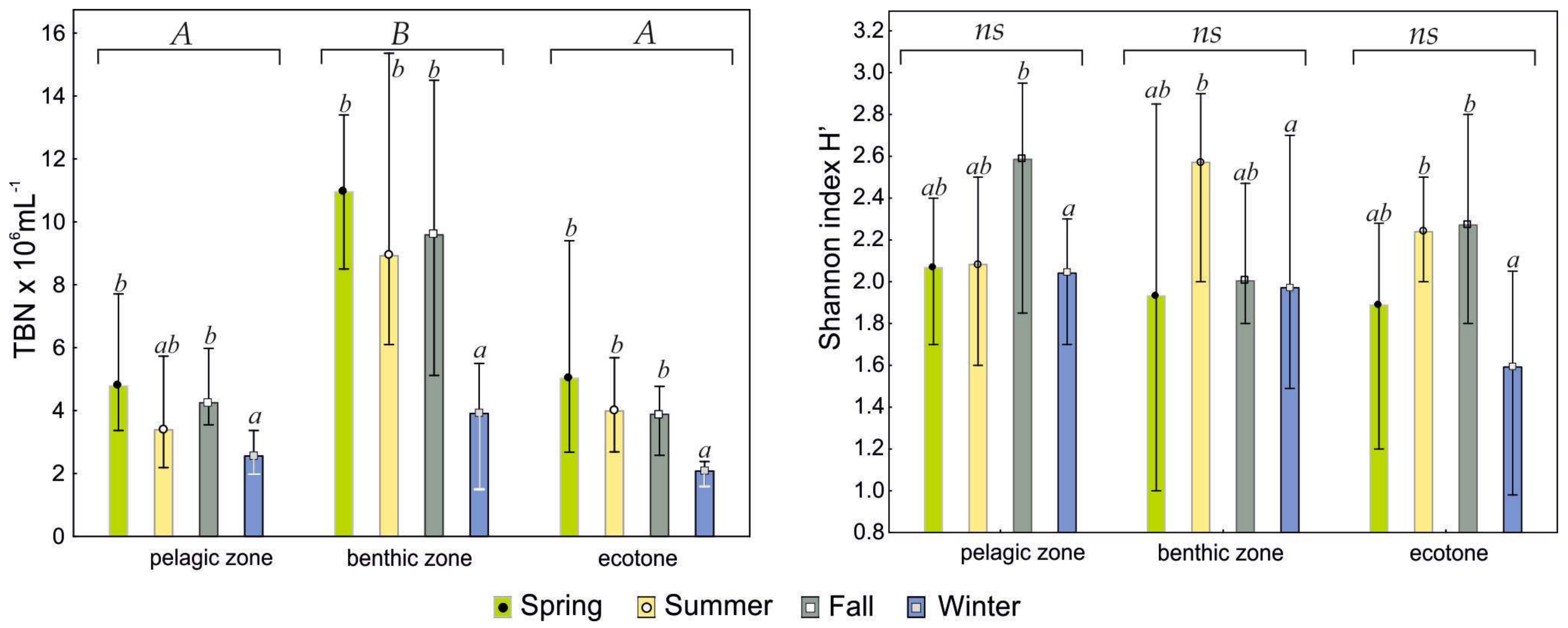
The average number of bacteria (TBN) stated in water of the studied peat bog lakes amounted to 5.3 × 106 mL−1. The highest TBN was recorded in the bottom layer (8.3 × 106 mL−1), and it ranged on average from 3.9 × 106 mL−1 in winter to 11.0 × 106 mL−1 in spring (Figure 3). The average TBNs noted for the benthic zone was statistically higher when compared to other two zones (Tukey’s test, post hoc, one-way ANOVA, p < 0.05). The pelagic (subsurface) and ecotone zones showed no statistical difference (3.7 × 106 mL−1) but showed a clear pattern of seasonal changes: maximum in spring and fall (4.9 and 4.1 × 106 mL−1, respectively) and minimum in winter (ca. 2.1 × 106 mL−1).
The lowest values of Shannon’s biodiversity index (H’) were also recorded in the ecotone zone in winter (1.0 to 2.05), but no typical pattern of seasonal changes was found for this parameter, unlike for TBN. A significantly higher values of biodiversity index (H’ = 2.60) were stated for the pelagic zone in fall, as well as for the benthic zone in summer (Tukey test, two-way ANOVA, p < 0.05), however, significantly lower values of the index (H’ = 1.6) appeared in winter months (Figure 3). No significant differences among three studied zones (Tukey’s test, post hoc, one-way ANOVA, p < 0.05) were detected as for H’. Seasonal pattern of the Pielou evenness index was presented Supplementary material (Figure S2).
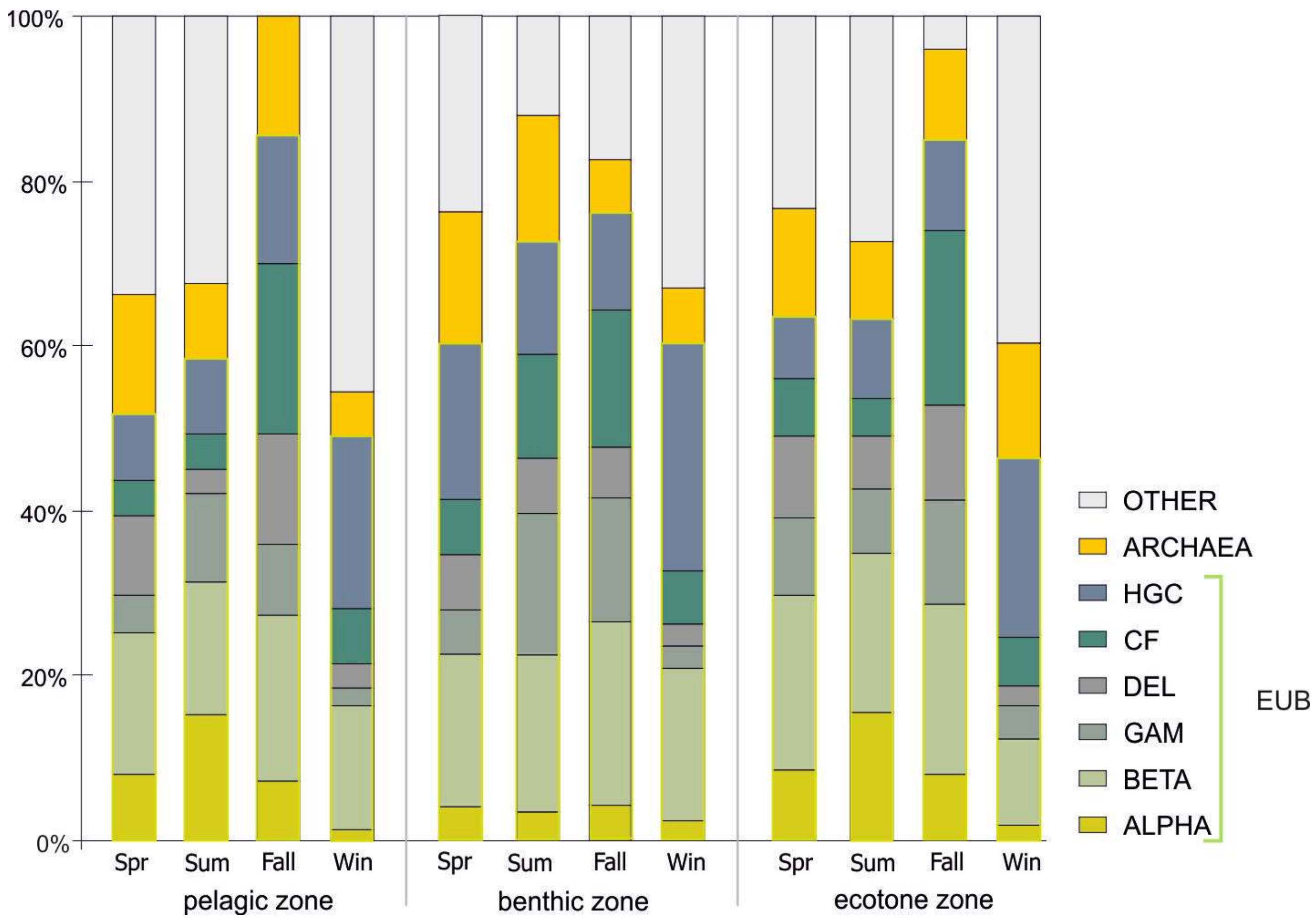
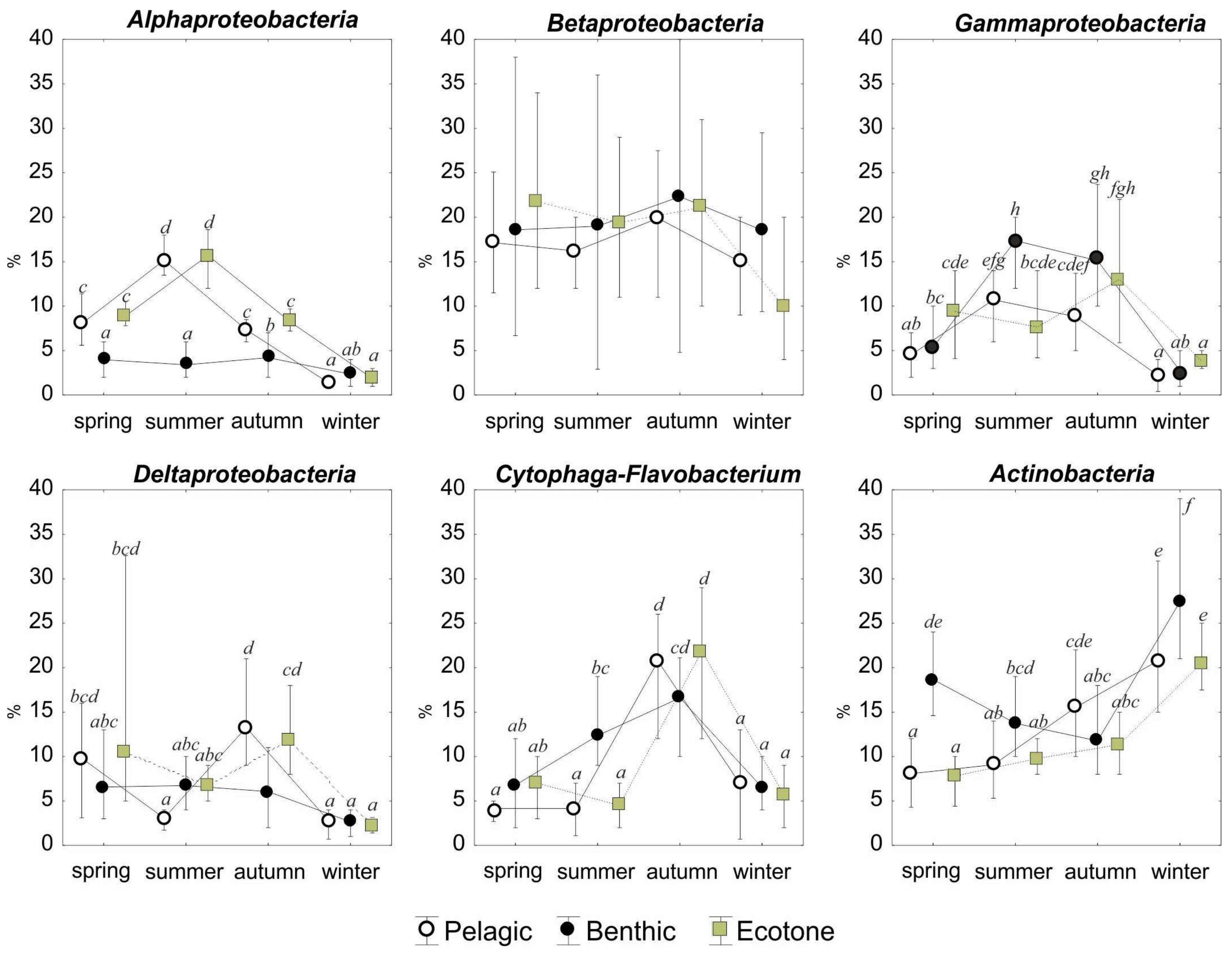
The relative abundance of Eubacteria in prokaryotic communities amounted on average to 64.5% (±15.42), and Archaea comprised as much as 11.3% (±4.78), while 24.3% were unidentified. The analysis of BCC demonstrated that, despite considerable differences between seasons, Betaproteobacteria constituted the largest class of bacteria and its share in the TBN amounted to 18.3% (±8.24) (Figure 4). Betaproteobacteria dominated regardless of a site or season, except for winter, when the share of Actinobacteria increased to 22.9%. Another abundant group of microorganisms, particularly often noted in the bottom, were Actinobacteria (15.5% ± 8.12 of TBN) and Gammaproteobacteria (8.3% ± 5.55 of TBN), (Figure 5). Actinobacteria were clearly more frequent during the spring and winter, while Gammaproteobacteria clearly filled the gap in other seasons, when water temperature was higher, i.e., in late summer and early fall.
The contribution of Alphaproteobacteria in the studied peat bog lakes amounted on average to 6.7% ± 4.8, of TBN. The group of bacteria is characterized by a specific pattern of seasonal changes for pelagic and ecotone zones. Its maximum (ca. 15% ± 1.56) was noted in summer and minimum (ca. 1.8% ± 0.6) in winter. In the bottom layer, Alphaproteobacteria constituted a small fraction of the community (<4.3%) throughout the whole study period with no clear pattern of seasonal changes (Figure 5).
Average abundances of Deltaproteobacteria and Cytophaga-Flavobacterium in peat bog lakes amounted to 6.8 ± 4.98 and 9.8% ± 7.03 of TBN, respectively. Their increased contributions in the bacteria community increased usually in fall in all the analyzed study sites and comprised 10.3% ± 4.59 and 19.7% ± 5.18, respectively.
Archaea in the microbial community showed different patterns for each sampling site (Figure 4). In the surface layer, two quantitative peaks were observed for this group of microorganisms: in spring and in fall, when their share amounted to 14.5% of the community. In the bottom in the first half of the year (spring and summer) their share did not drop below 10% in relation to all microorganisms stained with DAPI, and in other seasons its share was reduced on average by half. The number of Archaea was the most stable and relatively high (11.8%) in the ecotone zone comparing to pelagic and benthic zones. It is worth noticing that, except for the bottom, the lowest number of Archaea were recorded in summer (Figure 4).
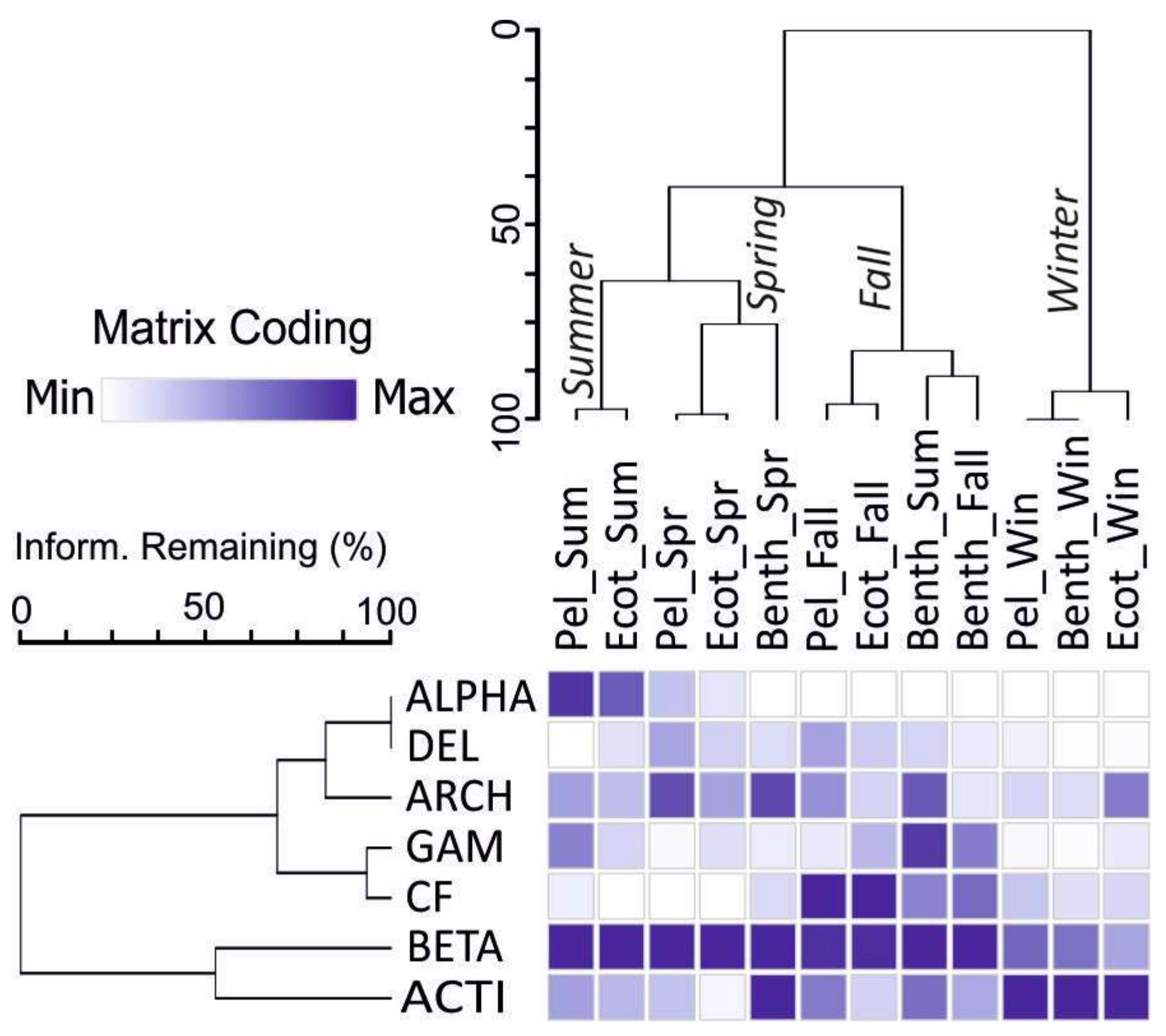
A heat map associated with the two-way cluster analysis allowed for visualization of the distribution of an individual bacteria phylum based on the relative population in samples (Figure 6). It can be seen that Betaproteobacteria dominate over the other bacteria groups regardless of habitat and season, except for winter.
3.3. Relationship between Physicochemical Parameters and Bacterial Community Composition
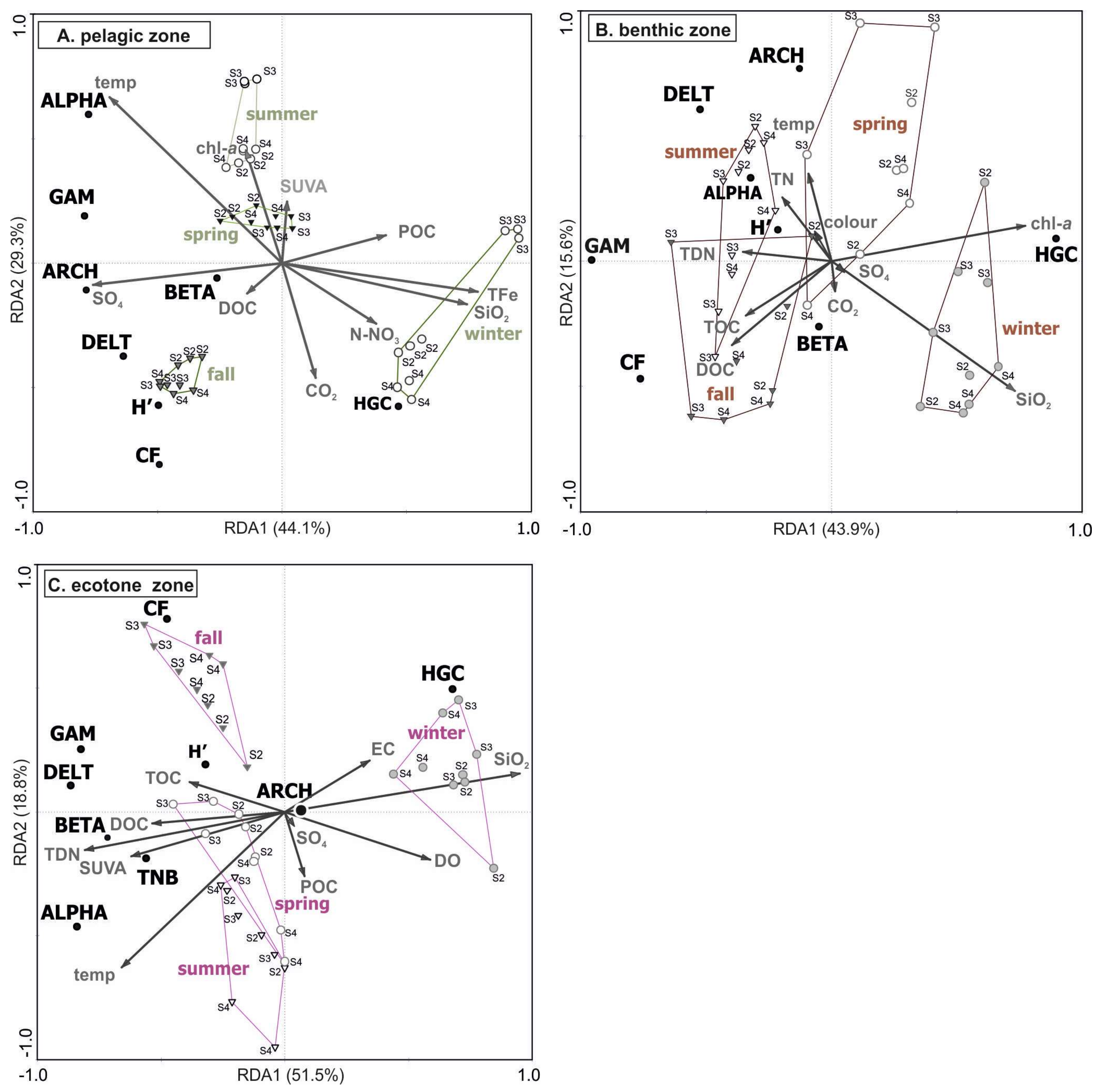
An RDA analysis was performed to examine the environmental factors determining the dynamics of different bacteria clades in the Suchar lakes. Regardless of a lake site (pelagic, benthic, or ecotonal) the two main factors (axes) explained more than 70% of the total variance.
Forward selection of explanatory (environmental) variables in the RDA model computed for the pelagic zone (Figure 7A) included nine out of 18 input variables (lambda (λA) significant at p < 0.003: water temperature (λA = 0.35), SO4 (λA = 0.17), SiO2 (λA = 0.10), N-NO3 (λA = 0.06), TFe (λA = 0.04), as well as DOC (λA = 0.04), POC (λA = 0.04), chl-a (λA = 0.04), and SUVA254 (λA = 0.03). The non-retained were redundant or did not increase the significance (e.g., pH). Most variables influencing the abundance of microbial communities were connected with the factor RDA1 displaying a significant gradient with temperature, SO4, and SIO2, while the factor RDA2 was related to organic carbon content and its aromaticity. In the well-sunlit pelagic zone, water temperature played an important role in determining the abundance of Alphaproteobacteria and Gammaproteobacteria while Actinobacteria showed an opposite character: their activity increased along with decreasing temperature gradients. Actinobacteria are well adapted to winter conditions when lower temperatures are accompanied by lower DOC levels. SO4 positively contributed to the abundance of Archaea. The RDA analysis also revealed that both TFe and SiO2 did not favor the bacteria abundance (in particular Archaea and Alpha-, Beta-, and Gammaproteobacteria), as their high concentrations in winter months.
Ten out of 18 input explanatory variables were retained as significant contributors to the RDA model for the benthic zone (Figure 7B). The most significant factor connected with RDA1 and displaying a positive gradient with the axis were SiO2 (λA = 0.28) and chl-a (λA = 0.15) concentrations. The latter contributed mostly to the presence of Actinobacteria at the benthic zone. The BCC was also significantly influenced by the availability of total and dissolved organic carbon. Conditional effects of TOC and DOC explained as much as λA = 0.06 and λA = 0.11 (p = 0.003) of variance, respectively. They both were found to be main drivers of Cytophaga-Flavobacteria abundance and, thus, factors contributing to the increased metabolic activity at the bottom zone.
The final RDA model for the ecotone zone accounted for 70.3% of the total variance in BCC composition and all canonical axes were significant (Monte Carlo test, p = 0.002). RDA model (Figure 7C) showed the role of the following factors displaying a positive gradient with RDA1: SIO2 (λA = 0.47), DO (λA = 0.19), and EC (λA = 0.09). A negative gradient with the RDA1 was shown by SUVA254 (λA = 0.10), TDN (λA = 0.02), TOC (λA = 0.05), and DOC (λA = 0.02). The organic carbon and nitrogen gradients determined the activity of major groups of Eubacteria (except for Actinobacteria), as well as their diversity index (H’-DGGE). None of the studied factors contributed significantly to the Archaea abundance in the ecotone zone. Temperature and POC in this zone were related to RDA2, and the variance explained by the factors amounted to λA = 0.11 and λA = 0.03, respectively.
Among the environmental variables, silica appeared to be an important factor contributing to the increase in the Actinobacteria number, regardless of the site. The analysis of SiO2 concentrations over the three-year study period showed a distinct fluctuation with late winter maximum SiO2 > 20 mg∙L−1, and summer and fall minimum SiO2 (<2 mg∙L−1). The increased concentrations of silica in late winter/early spring months is associated with the activity of diatoms, the predominant algae of the winter maximum, as light conditions improve, which brings oxygen saturation just below the ice cover. All the sampling sites observed lower values of silicate during winter months, perhaps due to the low temperature and sedimentation of suspended silica particles. The higher values were noticed during the winter season were due to the addition of turbid rain and snow-melted water from surrounding areas.
RDA performed for the parameters characterizing the BCC in relation to environmental variables demonstrated the crucial role of seasonality in all sites. The highest biodiversity expressed by the Shannon’s index (H’-DGGE) was noted particularly in the pelagic and ecotone zones during fall (H’ = 2.58 and H’ = 2.27, respectively) and increased with the availability of organic carbon (Figure 7A,C). An example of PCR-DGGE resolution for lake water samples is presented in Supplementary Materials Figure S1. Qualitative changes in the bacterial composition in the benthic zone were associated to the amount of total nitrogen (TDN, TN) as well as water temperature (Figure 7B).
In spite pH occurred a redundant variable in the RDA, correlation analysis preformed showed that acidity of water is an important factor limiting the presence of Betaproteobacteria. We have also found a statistically-significant negative correlation of this group of bacteria with pH as r = −0.492 (p ≤ 0.01; r not shown in figures). The strongest correlation was noted in the ecotone (r = −0.816; p ≤ 0.001), slightly weaker in the benthic zone (r = −0.464; p ≤ 0.01), and under the surface (r = −0.388; p ≤ 0.05). As the presented data are resulted from in situ measurements, but not controlled experiment, the stated relationships between the groups of microorganisms and the studied environmental explanatory variables may be affected by other factors.
4. Discussion
Our study revealed site-significant differences in the bacterial community structure in three peat bog lakes, suggesting that the bacterial community response to environmental conditions could be equally sensitive as seasonal variation. The studied lakes, like all dystrophic waters, are characterized by a low abundance of microorganisms, but for some, 5.1 and even 13 × 106 cells·mL−1 were reported [3,17,18]. The increased number of cells usually correlates with the depth as confirmed our study. Deeper water layers of peat bog lakes create favorable conditions for the growth of microorganisms because of the availability of DOC and nitrogen content (TN) from deposition down along the water column. According to Pankratov [41], the mean population density of bacteria under the mat built of Sphagnum moss is 3.9 × 108 cells·mL−1, but it usually increases with lake depth [42]. In addition, it has been observed that, regardless of the site location, the largest number of bacteria was recorded in spring, which may result from the inflow of microorganisms from the Sphagnum mat surrounding the lakes during the spring snowmelt. Moreover, such surface runoffs bring easily-assimilable organic matter that stimulates bacterial growth [43].
Studies on peatlands revealed that the bacterial community changes with depth, both in terms of community composition and size [41,44]. The diversity of microbes in peat bogs is influenced by biological factors, such as competition or predation, or changes in the specialization of communities caused by various abiotic factors [42]. Similar relationships determine the composition and diversity of bacterial communities in peat bog lakes. According to Hessen [45] the mean annual biomass of planktonic bacteria showed large variations both within and between lakes. The author reported that the lowest bacterial biomass was found in acidified lakes (7.8–12.1 μg C·L−1), and tended to increase with increasing water color (up to 44.1 μg C·L−1). Biodiversity indices determined based on the PCR-DGGE technique revealed no significant differences between the investigated habitats. Biodiversity calculated based on DGGE patterns, expressed by the Shannon’s H’ index, was the highest in fall in the ecotone and surface layers of the lakes and, in summer, just above the nutrient-rich bottom. Our analyses showed lower the H’ index values (from 1.0 to 2.9) regardless on a sampling site than the values reported in the clone library (3.53–3.69) for aquifer waters [46]. This supports the hypothesis stating that species diversity decreases in extreme conditions because only tolerant species survive in the community [47]. Peat bog lakes are extreme environments; additionally, the border zone (ecotone) is characterized by very variable hydrochemical conditions, which results in the low biodiversity of the bacterial community in peat bog lakes.
The literature reports on the strongest response of bacterial community to soil pH in peatland ecosystems [48]. Hessen [45] showed a strong positive correlation between bacterial biomass and humic content (r = 0.74, p < 0.001) and pH. Our study did not show such a strong response of the whole microbial community to this factor, but the density of dominant bacteria (Betaproteobacteria) was clearly correlated with changes in pH, particularly in the ecotone zone. The decrease in water pH was correlated with the dominance of this bacterial group and the simultaneous limitation in the growth of other microorganisms. Much of Betaproteobacteria are opportunistic bacteria, which easily adapt to changing environmental conditions [3,17,18], and this may explain their abundance during the whole study period in the three analyzed habitats.
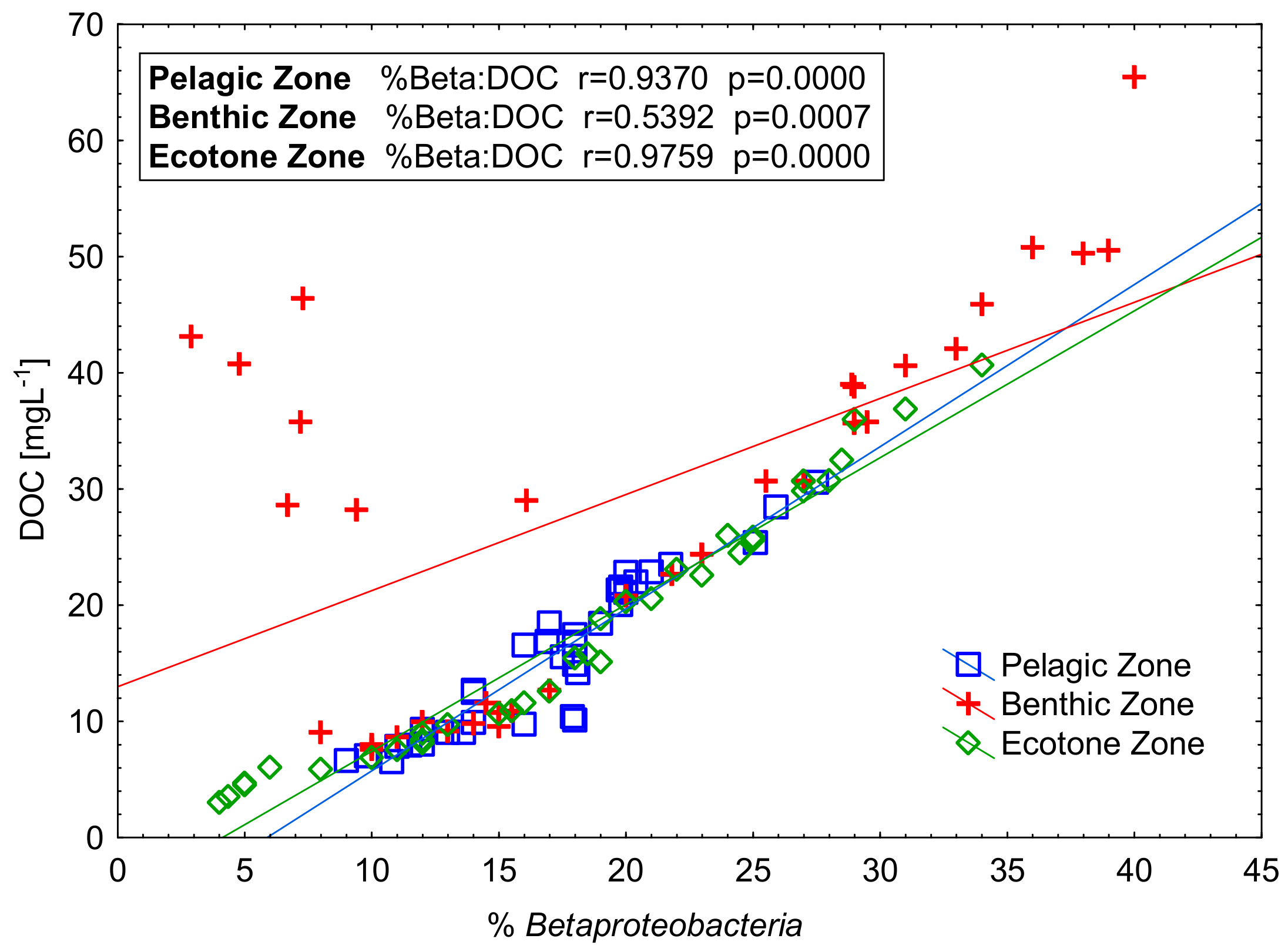
It is worth noting the clear positive linear correlation between Betaproteobacteria and DOC concentrations what is line with the observations reported by Bouvier [49]. The inflow of external allochthonic organic matter from the ecotone and its penetration down the water column through the lake layers less abundant in DOC was clearly correlated with the increased number of bacteria labelled with the Beta 42a probe.
Proteobacteria found in peat bog lakes most commonly belong to the Alpha- or Delta- classes. A strong similarity between the ecotone and surface layer of peat bog lakes was also noted in terms of the seasonal occurrence of other Proteobacteria. This was particularly evident for Alphaproteobacteria, which reached maximum abundance in spring and summer. In spring, some of them migrate to the lake from the adjacent area, where they are abundant in the Sphagnum mire surrounding peat bog lakes [46], and reach their quantitative maximum because of ideal conditions during summer months. Alphaproteobacteria are phototrophic bacteria preferring low-nutrient freshwater [50] and resistant to predation: their relative abundance increases in response to enhanced microeukaryote grazing [51]. Their share in the bacterial community is usually no greater than 5.5%, but because of perfect conditions, such as temperature, oxygen content, and exposure to sunlight, a three-fold increase in their share during summer was observed. Members of Alphaproteobacteria are generally not very abundant, suggesting that unknown mechanisms are keeping the abundance of the bacteria low in freshwater lakes. One potential mechanism may relate to the competitive ability of the Alphaproteobacteria for organic and inorganic substrates. Investigations suggested that the low availability of organic nutrients favors certain members of the Alphaproteobacteria [51].
Peat-inhabiting Deltaproteobacteria belong to phylogenetic lineages represented by the genera Syntrophobacter, Syntrophus, Smithella, Geobacter, and Anaeromyxobacter. Morales [52] found the Deltaproteobacteria to be the most frequent and probably the key group in bog ecology, because of their wide spectrum of ecological traits (e.g., sulphate reduction, iron reduction, or fermentation). This explains higher abundance of these bacteria in pelagic zone of the lakes with somewhat increased sulphate levels. Deltaproteobacteria, like the previously-discussed group of Proteobacteria, showed seasonal changes in the analyzed sites, except for the bottom, where their share in the bacterial community was stable, below 7% on average. This is a diversified group of microorganisms, including a significant number of sulphate- and sulphur-reducing bacteria, identified in marine and freshwater sediments, as well as in soils [53], but also obligate aerobes and predator cells feeding on other bacteria. It is possible that Deltaproteobacteria occur during fall, soon after the number of other bacteria, hosts to these pathogens, increases in oxygenated zones.
Gammaproteobacteria prefer nutrient-rich habitats [54] because they are typical r-strategists able to rapidly exploit the extra nutrients made available [55], which is reflected in their higher abundance (mean 15% of TBN) in the bottom layer during summer and fall. The greater proportion of this group (5–13%) has already been reported for peat bog lakes in comparison to other types of freshwater [3]. This is also confirmed by the present study.
Cytophaga-Flavobacterium are chemoorganotrophic, usually aerobic, and able to decompose chitin, cellulose, and pectin [56]. Involvement in the mineralization of complex macromolecules [57], more intensive during fall, corresponds with our findings indicating the maximum number of Cytophaga-Flavobacterium in this season.
Actinobacteria are able to decompose high molecular weight compounds and play a crucial role in mineralization [58,59]. Their significant share in communities of bog lakes has been reported before [58], and explained by the ability to assimilate compounds in these specific water bodies, as well as the transport of allochtonic bacteria from the surrounding mire. Members of this phylum can produce extracellular enzymes and have enzymatic capabilities comparable with those of fungi. They are known for their ability to degrade cellulose, lignin, and other complex biopolymers in different soil habitats, which enable them to survive in environments with low C availability [59]. Our results indicated that they can dominate the bacterial community at lower temperatures at low C content, which can be an analogue to certain arctic habitats [60]. Generally speaking, the freshwater lake Actinobacteria are free-living, open-water defence specialists, with possible photo- and heterotrophic energy generation lifestyles [51,61].
The phylum of Actinobacteria shows a clear exception to the variation in BCC over time (Figure 5). Warnecke [62] as well as Newton [19] suggest that the Actinobacteria does not constitute a transient component of lake communities originating from the soil in the catchment, but represents a unique pelagic freshwater lineage capable of sustaining growth in the lake. The dominance of Actinobacteria might be connected with a higher input of lignin-dominated spruce foliage. Actinobacteria seem to be involved in the process of cellulose degradation in acidic peat and are easily out-competed by Cytophaga-like bacteria under conditions of increased nitrogen availability [41]. The phylogenetic composition and potential role of bacteria participating in the degradation of Sphagnum-derived litter in ombrotrophic bogs is still largely unknown [41]. Whereas the Actinobacteria seem to be more or less uniform with regard to their metabolism and ecological requirements, Proteobacteria exhibit many different lifestyles and, therefore, the relative abundance of the main proteobacterial classes also varied between the lake sites. Some members of Proteobacteria are known to utilize easily available substrates quickly, as well as having the ability to use different compounds as electron donors [63].
Archeons, despite being the third domain of life, are still little known. They are present in all environments, from moderate to extreme and have recently increasing evidence reveals their widespread presence in various nonextreme environments, including soil, ocean, and freshwaters [64]. Two main archaeal phyla, Euryarchaeota and Crenarchaeota, are presently recognized. Recent phylogenetic studies suggest that the mesophilic Crenarchaeota form a third phylum, Thaumarchaeota [65]. Among Archaeas, an important group are methanogens [66], which in wetland ecosystems are responsible for emit approximately 3 to 7% of the global annual emission of greenhouse gas methane [67]. For this reason, peat-bog lakes seems to be ideal ecosystems to look for Archaea. A large proportion of them are found in anaerobic, cold, sulphide-rich aquatic habitats [68]. However, they do not prefer low temperatures, as under such conditions they only account for up to 5% of the bacterial community [60]. In our study, both in the surface and bottom layers, during the cold season in winter the share of Archaea was significantly lower in comparison to other seasons, when this group accounted on average for 10% to 16% of the total community. Archaea identified in the bottom layer did not represent the group of bacteria preferring anaerobic conditions, and were not involved in sulphate reduction and methanogenesis, in contrast to those usually found in bottom sediments [69]. The share of Archaea identified in our study decreased when the oxygen concentration in water was low. We found no significant correlation between water pH and the presence of Archaea, unlike other authors [70], which is clearly confirmed by our studies on acidic peat bog lakes.
5. Conclusions
The still insufficient knowledge about the microbial communities of peat bog lakes stimulates further research and field studies on these ecosystems. The use of recently-developed techniques based on molecular analysis helps in the study of bacterial community composition (BCC), which was previously very limited due to the fact that a considerable proportion of these bacteria are unable to grow on culture media. Peatland habitats play a significant environmental role. They can influence climate change, are rare not only in Europe, but worldwide, and may, due to their extensive character, be a source of new, interesting Bacteria and Archaea. Our study demonstrated that the acidic, shallow, and small lakes analyzed in the three seemingly different zones (pelagic, benthic, and ecotone), in fact, can be categorized only as two distinct habitats, which, despite their proximity, differ in qualitative and quantitative terms. The first habitat includes the bottom layer, and the second includes the ecotone and surface, where similar patterns of seasonal changes in bacterial composition and density are observed. The abundance of microorganisms has been proven for peat bog lakes, particularly for the bottom, which, despite apparently unfavorable conditions for other organisms, is an excellent zone for the proliferation of Bacteria and Archaea. The low pH and dark water color characteristic of these lakes do not directly influence the biodiversity or share of large groups of microorganisms in the community. A more important role is attributed to abiotic factors which regulate the microbial community in typical freshwater: temperature, levels of oxygen, nitrogen, and phosphorus, as well as easily-assimilable carbon (DOC). The quantitative and qualitative composition of microorganisms in peat bog lakes is also strongly influenced by the Sphagnum zone surrounding the lakes. The ecotone zone, as a source of allochthonic organic matter, is particularly important for the pelagic zone, which is naturally poor in autochthonic organic matter.
The results of our study have demonstrated that the peat bog lakes represent aquatic environments characterized by low bacterioplankton abundance and a site-specific, little-differentiated structure of the prokaryotic community. Our findings clarify the distribution patterns of the prokaryotic community composition in peat bog lakes, as well as evaluate the effects of environmental variable factors on prokaryotic community structures. Nevertheless, our findings do not fully confirm previous reports on the greater biodiversity within the ecotone zone. The quantitative and qualitative structure of Eubacteria seems to be a useful indicator of habitat conditions in peat-bog lakes. Our results indicate that this group of Prokaryotes does not differ significantly between the pelagial and ecotone zones. Considering the above, we have concluded that the similarity between pelagial and ecotone zones in peat-bog lakes distinguishes such lakes from other freshwater habitats.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/10/4/485/s1, Figure S1: An example of PCR-DGGE separation of 180 bp PCR fragment of V3 region of 16S rRNA gene, amplified with 338f-GC and 518r primers, visualized under UV light for bacterial communities of the water samples of chosen Suchar II, III, and IV lakes; samples 1–4 from Suchar II, 5–7 from Suchar III, 8–14 Suchar IV. The Shannon index values for these samples respectively: 2.19; 1.6; 2.47; 2.76; 2.28; 1.9; 1.3; 1.85; 2.46; 2.0; 1.75; 2.08; 0.7; 1.84, Figure S2: Seasonal and sites changes in Pielou index in peat-bog lakes. On the basis of Shannon Biodiversity Index, Pielou evenness index was calculated (Pielou, 1974) presenting the real diversity to maximal diversity ratio as a value between 0 and 1, Table S1: Oligonucleotide probes used in this work.
Acknowledgments
This study was supported by a grant within the scientific investigation project of the National Science Center (N N305 016940). Special thanks go to: the Management Board of Wigierski National Park and especially to Maciej Kamiński for allowing us to carry out the studies in WPN.
Author Contributions
S.L. conceived and designed the experiments; S.L. analyzed the data; A.Z.-B. contributed reagents and analysis tools; S.L. and K.G.-L. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mieczan, T.; Tarkowska-Kukuryk, M. Diurnal dynamics of the microbial loop in peatlands, structure, function and relationship to environmental parameters. Hydrobiologia 2013, 717, 189–201. [Google Scholar] [CrossRef]
- Cole, J.J.; Pace, M.L.; Carpenter, S.R.; Kitchell, J.F. Persistence of net heterotrophy in lakes during nutrient addition and food web manipulations. Limnol. Oceanogr. 2000, 45, 1718–1730. [Google Scholar] [CrossRef]
- Lew, S.; Koblížek, M.; Lew, M.; Medová, H.; Glińska-Lewczuk, K.; Owsianny, P. Seasonal changes of microbial communities in two shallow peat-bog lakes. Folia Microbiol. 2015, 60, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Tranvik, L.J. Dystrophy in Freshwater Systems. In Reference Module in Earth Systems and Environmental Sciences; On-Line: Current as of 25 November 2017; Elsevier: New York, NY, USA, 2014; ISBN 9780124095489. [Google Scholar] [CrossRef]
- Gudasz, C.; Bastviken, D.; Premke, K.; Steger, K.; Tranvik, L.J. Constrained microbial processing of allochthonous organic carbon in boreal lake sediments. Limnol. Oceanogr. 2012, 57, 163–175. [Google Scholar] [CrossRef]
- Grover, J.P. Resource competition and community structure in aquatic microorganisms: Experimental studies of algae and bacteria along a gradient of organic carbon to inorganic phosphorus supply. J. Plankton Res. 2000, 22, 1591–1610. [Google Scholar] [CrossRef]
- Yannarell, A.C.; Kent, A.D.; Lauster, G.H.; Kratz, T.K.; Triplett, E.W. Temporal patterns in bacterial communities in three temperate lakes of different trophic status. Microb. Ecol. 2003, 46, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, K.A.; Steven, F.; Findlay, R.; Neely, K.R. Priming in the microbial landscape: Periphytic algal stimulation of litter-associated microbial decomposers. Ecology 2014, 95, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Findlay, S.E.G.; Sinsabaugh, R.L.; Sobczak, W.V.; Hoostal, M. Metabolic and structural responses of hyporheic microbial communities to variations in supply of dissolved organic matter. Limnol. Oceanogr. 2003, 48, 1608–1617. [Google Scholar] [CrossRef]
- Amaral, V.; Graeber, D.; Calliari, D.; Alonso, C. Strong linkages between DOM optical properties and main clades of aquatic bacteria. Limnol. Oceanogr. 2016, 61, 906–918. [Google Scholar] [CrossRef]
- Von Wachenfeldt, E.; Sobek, S.; Bastviken, D.; Tranvik, L.J. Linking allochthonous dissolved organic matter and boreal lake sediment carbon sequestration: The role of light-mediated flocculation. Limnol. Oceanogr. 2008, 53, 2416–2426. [Google Scholar] [CrossRef]
- Wilkinson, G.M.; Pace, M.L.L.; Cole, J.J. Terrestrial dominance of organic matter in north temperate lakes. Glob. Biogeochem. Cycl. 2013, 27, 43–51. [Google Scholar] [CrossRef]
- Dedysh, S.N. Cultivating Uncultured Bacteria from Northern Wetlands: Knowledge Gained and Remaining Gaps. Front. Microbiol. 2011, 2, 184. [Google Scholar] [CrossRef] [PubMed]
- Roth, V.N.; Dittmar, T.; Gaupp, R.; Gleixner, G. Ecosystem-specific composition of dissolved organic matter. Vadose Zone J. 2014, 13, 7. [Google Scholar] [CrossRef]
- Ruiz-González, C.; Niño-García, J.P.; Lapierre, J.-F.; del Giorgio, P.A. The quality of organic matter shapes the functional biogeography of bacterioplankton across boreal freshwater ecosystems. Glob. Ecol. Biogeogr. 2015, 24, 1487–1498. [Google Scholar] [CrossRef]
- Arvola, L.; Eloranta, P.; Järvinen, M.; Keskitalo, J.; Holo-Painen, A.-L. Phytoplankton. In Limnology of Humic Waters; Keskitalo, J., Elo-Ranta, P., Eds.; Bachkhuys Publishers: Leiden, The Netherlands, 1999; pp. 137–171. [Google Scholar]
- Taipale, S.; Jones, R.I.; Tiirola, M. Vertical diversity of bacteria in an oxygen-stratified humic lake, evaluated using DNA and phospholipid analyses. Aquat. Microb. Ecol. 2009, 55, 1–16. [Google Scholar] [CrossRef]
- Burkert, U.F.; Warnecke, H.D.; Babenzien, E.; Zwirnmann, P.J. Members of a readily enriched-proteobacterial clade are common in surface waters of a humic lake. Appl. Environ. Microbiol. 2003, 68, 6550–6559. [Google Scholar] [CrossRef]
- Newton, R.J.; Kent, A.D.; Triplett, E.W.; McMahon, K.D. Microbial community dynamics in a humic lake: Differential persistence of common freshwater phylotypes. Environ. Microbiol. 2006, 8, 956–970. [Google Scholar] [CrossRef] [PubMed]
- Lindström, E.S. Bacterioplankton community composition in five lakes differing in trophic status and humic content. Microb. Ecol. 2000, 40, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Benatti, C.T.; Tavares, C.R.G.; Lenzi, E. Sulfate removal from waste chemicals by precipitation. J. Environ. Manag. 2009, 90, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Sanders, R.; Tappin, A.D.; Worsfold, P.J.; Achterberg, E.P. Simultaneous determination of dissolved organic carbon and total dissolved nitrogen on a coupled high-temperature combustion total organic carbon-nitrogen chemiluminescence detection HTC TOC-NCD system. J. Autom. Methods Manag. Chem. 2005, 2005, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Paul, J.H.; Cazares, L.; Thurmond, J. Amplification of the rbcL gene from dissolved and particulate DNA from aquatic environments. Appl. Environ. Microbiol. 1990, 56, 1963–1966. [Google Scholar] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Aquat. Microb. Ecol. 1993, 59, 695–700. [Google Scholar]
- Ziembińska, A.; Ciesielski, S.; Miksch, K. Ammonia oxidizing bacteria community in activated sludge monitored by denaturing gradient gel electrophoresis DGGE. J. Gen. Appl. Microbiol. 2009, 55, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, Y.; Wang, Y.H.; Liu, K.; Tong, L. Bacterial community structure and diversity during establishment of an anaerobic bioreactor to treat swine wastewater. Water Sci. Technol. 2010, 1, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Glöckner, F.O.; Amann, R.; Alfreider, A.; Pernthaler, J.; Psenner, R.; Trebesius, R.; Schleifer, K.H. An in situ hybridization protocol for detection and identyfication of planctonic bacteria. Syst. Appl. Microbiol. 1996, 19, 403–406. [Google Scholar] [CrossRef]
- Pernthaler, A.; Pernthaler, J.; Amann, R. Fluorescent in situ hybridization, FISH, with rRNA-targeted oligonucleotide probes, review. Methods Microbiol. 2001, 30, 207–226. [Google Scholar] [CrossRef]
- Stoecker, K.; Dorninger, C.; Daims, H.; Wagner, M. Double Labeling of Oligonucleotide Probes for Fluorescence In Situ Hybridization, DOPE-FISH, Improves Signal Intensity and Increases rRNA Accessibility. Appl. Environ. Microb. 2010, 76, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria; development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Stahl, D.A.; Amann, R. Development and Application of Nucleic Acid Probes. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 205–248. [Google Scholar]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria, problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Loy, A.; Lehner, A.; Lee, N.; Adamczyk, J.; Meier, H.; Ernst, J.; Schleifer, K.-H.; Wagner, M. Oligonucleotide microarray for 16S rRNA gene-based detection of all recognized lineages of sulfate-reducing prokaryotes in the environment. Appl. Environ. Microbiol. 2002, 68, 5064–5081. [Google Scholar] [CrossRef] [PubMed]
- Roller, C.; Wagner, M.; Amann, R.; Ludwig, W.; Schleifer, K.H. In situ probing of Gram-positive bacteria with high DNA G+C content using 23S rRNA-targeted oligonucleotides. Microbiology 1994, 140, 2849–2858. [Google Scholar] [CrossRef] [PubMed]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization with rRNA-targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Ter Braak, C.J.F.; Śmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide, Software for Canonical Community Ordination; Version 4.5; Microcomputer Power: Ithaca, NY, USA, 2002; p. 500. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using Canoco; Cambridge University Press: Cambridge, UK, 2003; p. 269. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. In Multivariate Analysis of Ecological Data; Version 6; MjM Software: Gleneden Beach, OR, USA, 2010. [Google Scholar]
- Glinska-Lewczuk, K. Effect of Land Use and Lake Presence on Chemical Diversity of the Łyna River System. Pol. J. Environ. Stud. 2006, 15, 259–269. [Google Scholar]
- Pankratov, T.A.; Dedysh, S.N.; Zavarzin, G.A. The Leading Role of Actinobacteria in Aerobic Cellulose Degradation in Sphagnum Peat Bogs. Microbiology 2006, 410, 564–567. [Google Scholar] [CrossRef]
- Andersen, R.; Chapman, S.J.; Artz, R.R.E. Microbial communities in natural and disturbed peatlands: A review. Soil Biol. Biochem. 2013, 57, 979–994. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Pankratov, T.A.; Belova, S.E.; Kulichevskaya, I.S.; Liesack, W. Phylogenetic analysis and in situ identification of bacteria community composition in an acidic Sphagnum peat bog. Appl. Environ. Microbiol. 2006, 72, 2110–2117. [Google Scholar] [CrossRef] [PubMed]
- Sieburth, J.M. Microbiological and organic–chemical processes in the surface and mixed layers. In Air–Sea Exchange of Gases and Particles; Liss, P.S., Slinn, W.G.N., Eds.; Riedel Publishers Co.: Hingham, MA, USA, 1983; pp. 121–172. [Google Scholar]
- Hessen, D.O. The relation between bacterial carbon and dissolved humic compounds in oligotrophic lakes. FEMS Microbiol. Ecol. 1985, 1, 215–223. [Google Scholar] [CrossRef]
- Koizumi, Y.; Kojima, H.; Fukui, M. Characterization of depth-related microbial community structure in lake sediment by denaturing gradient gel electrophoresis of amplifed 16S rDNA and reversely transcribed 16S rRNA fragments. FEMS Microb. Ecol. 2003, 46, 147–157. [Google Scholar] [CrossRef]
- Goździejewska, A.; Glińska-Lewczuk, K.; Obolewski, K.; Grzybowski, M.; Kujawa, R.; Lew, S.; Grabowska, M. Effects of lateral connectivity on zooplankton community structure in floodplain lakes. Hydrobiologia 2016, 774, 7. [Google Scholar] [CrossRef]
- Hartman, W.H.; Richardson, C.J.; Vilgalys, R.; Bruland, G.L. Environmental and anthropogenic controls over bacterial communities in wetland soils. Proc. Natl. Acad. Sci. USA 2008, 105, 17842–17847. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, T.; del Giorgio, P.A. Compositional changes in freeliving bacterial communities along a salinity gradient in two temperate estuaries. Limnol. Oceanogr. 2002, 47, 453–470. [Google Scholar] [CrossRef]
- Imhoff, J.F. The phototrophic Alpha-proteobacteria. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schlbifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 541–564. ISBN 978-0-387-30745-9. [Google Scholar]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S.A. Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed]
- Morales, S.E.; Mouser, P.J.; Ward, N.; Hudman, S.P.; Gotelli, N.J.; Ross, D.S.; Lewis, T.A. Comparison of bacterial communities in New England Sphagnum bogs using terminal restriction fragment length polymorphism. Microb. Ecol. 2006, 52, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Mussmann, M.; Ishii, K.; Rabus, R.; Amann, R. Diversity and vertical distribution of cultured and uncultured Deltaproteobacteria in an intertidal mud flat of the Wadden Sea. Environ. Microb. 2005, 7, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Pinhassi, J.; Berman, T. Differential growth response of colony-forming α- and γ-Proteobacteria in dilution culture and nutrient addition experiments from Lake Kinneret, Israel, the Eastern Mediterranean Sea, and the Gulf of Eilat. Appl. Environ. Microbiol. 2003, 69, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, R.; Callieri, C.; Balseiro, E.; Modenutti, B. Susceptibility of bacterioplankton to nutrient enrichment of oligotrophic and ultraoligotrophic lake waters. J. Limnol. 2008, 67, 120–127. [Google Scholar] [CrossRef]
- Kirchman, D. The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microb. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Fuchs, B.M.; Amann, R. Bacterioplankton compositions of lakes and oceans, a first comparison based on fluorescence in situ hybridization. Appl. Environ. Microbiol. 1999, 65, 3721–3726. [Google Scholar] [PubMed]
- Buck, U.; Grossaet, H.-P.; Amann, R.; Pernthaler, J. Substrate incorporation patterns of bacterioplankton populations in stratified and mixed waters of a humic lake. Environ. Microbiol. 2009, 11, 1854–1865. [Google Scholar] [CrossRef] [PubMed]
- Rheims, H.; Felske, A.; Seufert, S.; Stackebrandt, M. Molecular monitoring of an uncultured group of the class Actinobacteria in two terrestrial environments. J. Microbiol. Methods 1999, 36, 65–75. [Google Scholar] [CrossRef]
- Høj, L.; Olsen, R.A.; Torsvik, V.L. Effects of temperature on the diversity and community structure of known methanogenic groups and other archaea in high Arctic peat. ISME J. 2008, 2, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Koblizek, M. Ecology of aerobic anoxygenic phototrophs in aquatic environments. FEMS Microbiol. Rev. 2015, 39, 854–870. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, F.; Amann, R.; Pernthaler, J. Actinobacterial 16S rRNA genes from freshwater habitats cluster in four distinct lineages. Environ. Microbiol. 2004, 6, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, Z.; Bárta, J. Microbial community composition and in silico predicted metabolic potential reflect biogeochemical gradients between distinct peatland types. FEMS Microbiol. Ecol. 2014, 90, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, F.; Kong, Z.; Yin, J.; Kou, W.; Wu, L.; Ge, G. The Distribution Pattern of Sediment Archaea Community of the Poyang Lake, the Largest Freshwater Lake in China. Archaea 2016, 2016, 9278929. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, M.; Montonen, L.; Münster, U.; Jurgens, G. Diversity and function of archaea in freshwater habitats. Curr. Trends Microbiol. 2008, 4, 1–89. [Google Scholar]
- Juottonen, H. Archaea, Bacteria, and Methane Production along Environmental Gradients in Fens and Bogs. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2008; p. 48. [Google Scholar]
- Horn, M.A.; Matthies, C.; Kusel, K.; Schramm, A.; Drake, H.L. Hydrogenotrophic methanogenesis by moderately acid-tolerant methanogens of a methane-emitting acidic peat. Appl. Environ. Microbiol. 2003, 69, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Llirós, M.; Casamayos, E.O.; Borrego, C. High archaeal richness in the water column of a freshwater sulfurous karstic lake along an inter-annual study. FEMS Microbiol. Ecol. 2008, 66, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, J.I.K.; Eckert, W.; Conrad, R. Community structure of Archaea and Bacteria in a profundal lake sediment Lake Kinneret, Israel. Syst. Appl. Microbiol. 2007, 30, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, P.; Sterngren, A.E.; Rousk, J. Archaeal abundance across a pH gradient in an Arable Soil and its relationship to bacterial and fungal growth rates. Appl. Environ. Microbiol. 2012, 78, 5906–5911. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Location of the study area and aerial view on the Suchar lakes II, III, and IV with main morphometric features.
Figure 1.
Location of the study area and aerial view on the Suchar lakes II, III, and IV with main morphometric features.

Figure 2.
Crossection of a peat bog lake with the ecotone zone surrounding the water body. White arrows indicate sampling sites.
Figure 2.
Crossection of a peat bog lake with the ecotone zone surrounding the water body. White arrows indicate sampling sites.

Figure 3.
Seasonal differences in abundances of bacteria (TBN) (left graph) and bacteria diversity expressed Shannon index (H’) (right graph) in the studied parts of the peat bog lakes. The heights of bars denote means for lakes and the error bars represent standard deviations. Means that do not share superscripts differ by p < 0.05 according to the Tukey’s test (post hoc, two-way ANOVA). The homogeneous subsets output, expressed by combination of ab superscripts, shows groups of means that do not differ significantly from each other. Statistical differences between groups of means for sites been indicated with capital letters A and B above the bars; ns—no statistical significance among H’ in seasons.
Figure 3.
Seasonal differences in abundances of bacteria (TBN) (left graph) and bacteria diversity expressed Shannon index (H’) (right graph) in the studied parts of the peat bog lakes. The heights of bars denote means for lakes and the error bars represent standard deviations. Means that do not share superscripts differ by p < 0.05 according to the Tukey’s test (post hoc, two-way ANOVA). The homogeneous subsets output, expressed by combination of ab superscripts, shows groups of means that do not differ significantly from each other. Statistical differences between groups of means for sites been indicated with capital letters A and B above the bars; ns—no statistical significance among H’ in seasons.

Figure 4.
Changes in the taxonomic composition of bacterial communities in the Suchar lakes II, III, and IV Sphagnum mat-surrounded peat bog lakes.
Figure 4.
Changes in the taxonomic composition of bacterial communities in the Suchar lakes II, III, and IV Sphagnum mat-surrounded peat bog lakes.

Figure 5.
Seasonal changes in the Eubacteria for the studied Suchar lakes. Values are expressed as the percentage of hybridized cell counts of TBN of DAPI-stained cells. Error bars represent standard errors. Means that do not share superscripts differ by p < 0.05 according to the Tukey’s test (post hoc, two-way ANOVA). The combination of superscript letters shows groups of means that do not differ significantly from each other.
Figure 5.
Seasonal changes in the Eubacteria for the studied Suchar lakes. Values are expressed as the percentage of hybridized cell counts of TBN of DAPI-stained cells. Error bars represent standard errors. Means that do not share superscripts differ by p < 0.05 according to the Tukey’s test (post hoc, two-way ANOVA). The combination of superscript letters shows groups of means that do not differ significantly from each other.

Figure 6.
Two way cluster analysis exposing relationship between clusters of Eubacteria-Archaea and seasons and sites. Heat map colors indicate minimum (white) to maximum (navy blue) abundances in samples, wherein colors are scaled by a relative abundance in a column.
Figure 6.
Two way cluster analysis exposing relationship between clusters of Eubacteria-Archaea and seasons and sites. Heat map colors indicate minimum (white) to maximum (navy blue) abundances in samples, wherein colors are scaled by a relative abundance in a column.

Figure 7.
RDA biplots of physiochemical and microbiological parameters for the pelagic (A), benthic (B), and ecotone zones (C). Environmental variables are represented by arrows that approximately point towards the factor direction of maximum variation. The length of an arrow is proportional to the importance of that variable in the ordination. The angles between response (bacteria descriptors) and explanatory (environmental) variables in the RDA biplots reflect their correlations. Groups of samples represented a given season is marked in color.
Figure 7.
RDA biplots of physiochemical and microbiological parameters for the pelagic (A), benthic (B), and ecotone zones (C). Environmental variables are represented by arrows that approximately point towards the factor direction of maximum variation. The length of an arrow is proportional to the importance of that variable in the ordination. The angles between response (bacteria descriptors) and explanatory (environmental) variables in the RDA biplots reflect their correlations. Groups of samples represented a given season is marked in color.

Figure 8.
The relationship between participation in bacterial community Betaproteobacteria and the content of dissolved organic carbon (DOC) in the zones of examined peat-bog lakes.
Figure 8.
The relationship between participation in bacterial community Betaproteobacteria and the content of dissolved organic carbon (DOC) in the zones of examined peat-bog lakes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Water quality parameters of the studied peat bog lakes. Values represent means ± standard deviation of measurements for three lakes in four seasons over a period of three years. Means bearing different superscripts (a, b) differ significantly by the Tukey’s test (post-hoc, ANOVA; p ≤ 0.05); pH—median, N—number of samples.
Table 1.
Water quality parameters of the studied peat bog lakes. Values represent means ± standard deviation of measurements for three lakes in four seasons over a period of three years. Means bearing different superscripts (a, b) differ significantly by the Tukey’s test (post-hoc, ANOVA; p ≤ 0.05); pH—median, N—number of samples.
| Parameter | N | pH | Temp. | EC | DO | Chl-a | Color | N-NO3 | N-NH4 | TDN | TN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Site/Unit | - | °C | µS cm−1 | mg L−1 | µg L−1 | mg PtL−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | |
| Pelagic Zone | 36 | 4.46 | 12.47 ± 7.66 b | 26.76 ± 7.97 a | 8.92 ± 2.60 a | 32.04 ± 40.64 a | 148 ± 74 a | 0.16 b ± 0.16 | 0.03 ± 0.02 a | 0.58 ± 0.16 a | 0.74 ± 0.21 a |
| Benthic Zone | 36 | 4.36 | 6.20 ± 3.09 a | 37.26 ± 9.52 b | 2.21 ± 1.72 a | 108.53 ± 44.92 b | 390 ± 328 b | 0.07 ± 0.06 a | 0.61 ± 0.58 b | 1.88 ± 1.01 b | 2.41 ± 1.50 b |
| Ecotone Zone | 36 | 4.33 | 12.18 ± 7.06 b | 26.65 ± 5.91 a | 7.34 ± 2.17 b | 28.05 ± 14.47 a | 151 ± 91 a | 0.05 ± 0.03 a | 0.08 ± 0.05 a | 0.65 ± 0.24 a | 1.00 ± 0.49 a |
| Total | 108 | 4.38 | 10.28 ± 6.86 | 30.22 ± 9.32 | 6.16 ± 3.61 | 45.74 ± 46.01 | 230 ± 229 | 0.10 ± 0.11 | 0.24 ± 0.43 | 1.03 ± 0.85 | 1.38 ± 1.17 |
| Site | P-PO4 | TP | TOP | SUVA254 | TOC | DOC | POC | TFe | SO4 | SiO2 | |
| /Unit | mg L−1 | mg L−1 | mg L−1 | L mg−1 m−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | |
| Pelagic Zone | 36 | 0.03 ± 0.03 a | 0.05 ± 0.02 a | 0.03 ± 0.02 a | 3.84 ± 0.48 | 18.15 ± 6.88 a | 15.60 ± 6.58 a | 2.49 ± 0.89 a | 0.33 ± 0.28 a | 1.12 ± 0.50 | 4.47 ± 7.43 |
| Benthic Zone | 36 | 0.05 ± 0.04 b | 0.11 ± 0.05 b | 0.06 ± 0.02 c | 4.20 ± 0.42 | 33.58 ± 18.25 b | 29.22 ± 16.68 b | 4.58 ± 2.63 b | 0.62 ± 0.28 b | 1.36 ± 0.70 | 8.08 ± 9.00 |
| Ecotone Zone | 36 | 0.02 ± 0.00 a | 0.06 ± 0.03 a | 0.05 ± 0.03b | 3.67 ± 0.79 | 18.48 ± 9.00 a | 17.60 ± 10.69 a | 2.29 ± 1.53 a | 0.30 ± 0.19 a | 1.40 ± 0.54 | 6.21 ± 8.84 |
| Total | 108 | 0.03 ± 0.03 | 0.08 ± 0.04 | 0.05 ± 0.03 | 3.91 ± 0.62 | 23.40 ± 14.26 | 20.81 ± 13.38 | 3.12 ± 2.09 | 0.42 ± 0.29 | 1.30 ± 0.59 | 6.82 ± 8.50 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lew, S.; Glińska-Lewczuk, K.; Ziembińska-Buczyńska, A. Prokaryotic Community Composition Affected by Seasonal Changes in Physicochemical Properties of Water in Peat Bog Lakes. Water 2018, 10, 485. https://doi.org/10.3390/w10040485
AMA Style
Lew S, Glińska-Lewczuk K, Ziembińska-Buczyńska A. Prokaryotic Community Composition Affected by Seasonal Changes in Physicochemical Properties of Water in Peat Bog Lakes. Water. 2018; 10(4):485. https://doi.org/10.3390/w10040485
Chicago/Turabian StyleLew, Sylwia, Katarzyna Glińska-Lewczuk, and Aleksandra Ziembińska-Buczyńska. 2018. "Prokaryotic Community Composition Affected by Seasonal Changes in Physicochemical Properties of Water in Peat Bog Lakes" Water 10, no. 4: 485. https://doi.org/10.3390/w10040485
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.